
วิวัฒนาการของตา
| ส่วนหนึ่งของชุดบทความเรื่อง |
| ชีววิทยาวิวัฒนาการ |
|---|
 |
วิวัฒนาการของตา (อังกฤษ: evolution of the eye) เป็นประเด็นการศึกษาที่ดึงดูดความสนใจ เพราะเป็นตัวอย่างพิเศษที่แสดงวิวัฒนาการเบนเข้าของอวัยวะที่สัตว์กลุ่มต่าง ๆ มากมายมี คือตาที่ซับซ้อนและทำให้สามารถมองเห็นได้วิวัฒนาการเกิดขึ้นอย่างเป็นอิสระกว่า 50-100 ครั้ง[1]
ตาที่ซับซ้อนดูเหมือนจะวิวัฒนาการขึ้นภายในไม่กี่ล้านปีในช่วง Cambrian explosion (เหตุการณ์ระเบิดสิ่งมีชีวิตยุคแคมเบรียน) ที่สิ่งมีชีวิตได้เกิดวิวัฒนาการอย่างรวดเร็ว หลักฐานว่าตาได้วิวัฒนาการขึ้นก่อนยุคแคมเบรียนยังไม่มี แต่มีอย่างหลากหลายในชั้นหิน/สิ่งทับถม Burgess shale (ในเทือกเขาร็อกกีของประเทศแคนาดา) ในกลางยุคแคมเบรียน และในหมวดหิน Emu Bay Shale ในออสเตรเลียซึ่งเก่าแก่กว่าเล็กน้อย[2]
ตาได้ปรับตัวอย่างหลายหลากตามความจำเป็นของสัตว์ ความต่างกันรวมทั้งความชัด (visual acuity) พิสัยความยาวคลื่นแสงที่สามารถเห็น ความไวในแสงสลัว ๆ สมรรถภาพในการตรวจจับการเคลื่อนไหวหรือการแยกแยะวัตถุ และการเห็นเป็นสี
ประวัติงานวิจัย[แก้]

ในปี 2345 นักปรัชญา William Paley ได้เรียกตาว่า เป็นสิ่งหัศจรรย์ในการสร้างโลก ส่วนชาลส์ ดาร์วินเองได้เขียนในหนังสือ Origin of Species ว่า วิวัฒนาการของตาผ่านกระบวนการคัดเลือกโดยธรรมชาติอาจจะดูเหมือนเรื่องเหลวไหลที่สุด แต่เขาก็อธิบายต่อไปว่า แม้จะจินตนาการได้ยาก แต่ก็เป็นไปได้อย่างแน่นอน คือ
..ถ้าการเปลี่ยนแปลงจากตาง่าย ๆ และไม่สมบูรณ์กลายเป็นตาที่ซับซ้อนและสมบูรณ์สามารถแสดงได้ว่ามี และการเปลี่ยนแปลงแต่ละอย่างก็มีประโยชน์ต่อผู้ที่มี ซึ่งก็จะต้องเป็นเช่นนี้อย่างแน่นอน ยิ่งกว่านั้น ถ้าตาต่าง ๆ กันไป และความต่างกันสามารถสืบทอดต่อไปได้ ซึ่งเช่นกัน ต้องเป็นอย่างแน่นอน และความหลายหลากเช่นนี้มีประโยชน์ต่อสัตว์ใด ๆ ที่ปัจจัยต่าง ๆ ในชีวิตกำลังแปรไป ความยากลำบากในการเชื่อว่าตาที่ซับซ้อนและสมบูรณ์สามารถเกิดขึ้นได้ผ่านการคัดเลือกโดยธรรมชาติโดยที่เราจินตนาการไม่ได้ ก็ไม่ควรพิจารณาว่า ล้มล้างทฤษฎี
— ชาลส์ ดาร์วิน ในหนังสือ Origin of Species พ.ศ. 2402[3]
เขาได้เสนอการเปลี่ยนแปลงจาก "ประสาทตาที่เพียงฉาบด้วยสารรงควัตถุโดยไร้กลไกอย่างอื่น ๆ" กลายเป็น "ตามที่สมบูรณ์ค่อนข้างสูง" และให้ตัวอย่างของการเปลี่ยนแปลงระยะต่าง ๆ ทางวิวัฒนาการที่มีอยู่จริง ๆ[3] ข้อเสนอของดาร์วินไม่นานก็ได้หลักฐานว่าถูกต้อง โดยงานวิจัยในปัจจุบันกำลังตรวจสอบกลไกทางพันธุกรรมที่เป็นเหตุของพัฒนาการและวิวัฒนาการของตา[4]
นักชีววิทยา D.E. Nilsson ได้เสนอระยะทั่วไป 4 ระยะในวิวัฒนาการของตาของสัตว์มีกระดูกสันหลังเริ่มตั้งแต่การเกิดเซลล์รับแสงแผ่นหนึ่ง[5] Nilsson และ Pelger ได้พิมพ์ผลงานคลาสสิกที่แสดงว่า จะใช้เวลากี่ชั่วยุคเพื่อวิวัฒนาการให้ได้ตาที่ซับซ้อนเช่นในสัตว์มีกระดูกสันหลัง[6] ส่วนนักชีววิทยา G.C. Young ได้ใช้หลักฐานซากดึกดำบรรพ์เพื่ออนุมานข้อสรุปต่าง ๆ ทางวิวัฒนาการ อาศัยโครงสร้างของเบ้าตาและช่องต่าง ๆ ในกะโหลกดึกดำบรรพ์ที่เส้นเลือดและเส้นประสาทจะวิ่งผ่าน[7] งานเหล่านี้ล้วนเพิ่มหลักฐานสนับสนุนทฤษฎีของดาร์วิน
อัตราการวิวัฒนาการ[แก้]
ซากดึกดำบรรพ์ที่แสดงหลักฐานเกี่ยวกับตาแรกสุด มาจากต้นยุคแคมเบรียนราว 540 ล้านปีก่อน[8] ซึ่งเป็นยุคที่เกิดวิวัฒนาการอย่างรวดเร็ว จึงได้ชื่อว่า "Cambrian explosion" (เหตุการณ์ระเบิดสิ่งมีชีวิตยุคแคมเบรียน) หนึ่งในบรรดาสมมติฐานเกี่ยวกับเหตุของความหลากหลายที่เกิดขึ้น ก็คือ ทฤษฎี "Light Switch" ของ ดร. แอนดรูว์ ปาร์เกอร์ ซึ่งเสนอว่า วิวัฒนาการของตาได้จุดชนวนการแข่งขันในทางอาวุธ ซึ่งทำให้เกิดวิวัฒนาการอย่างรวดเร็ว[9] ก่อนหน้ายุคนี้ สิ่งมีชีวิตอาจไวแสง แต่ก็ไม่ใช่เพื่อการเคลื่อนไหวอย่างรวดเร็วและการหาทิศทางโดยใช้ตา
การประมาณอัตราวิวัฒนาการของตาเป็นเรื่องยากเพราะซากดึกดำบรรพ์โดยเฉพาะจากยุคแคมเบรียนต้น ๆ มีน้อยมาก ดังนั้น วิวัฒนาการจากเซลล์รับแสงเป็นแผ่นกลม ๆ จนเกิดตาที่ใช้งานได้อย่างสมบูรณ์ จึงประเมินอาศัยอัตราการกลายพันธุ์ ประโยชน์ที่สัตว์ได้โดยเปรียบเทียบ และการคัดเลือกโดยธรรมชาติ อาศัยการคำนวณซึ่งมองในแง่ร้ายเพราะประเมินเวลาเกินอย่างสม่ำเสมอสำหรับเวลาในแต่ละระยะ ๆ และอาศัยการกำหนดชั่วยุคสัตว์ที่ 1 ปี ซึ่งสามัญในสัตว์เล็ก ๆ นักวิทยาศาสตร์ได้เสนอว่า มันจะใช้เวลาน้อยกว่า 364,000 ปีเพื่อวิวัฒนาการเกิดตาในสัตว์มีกระดูกสันหลังจากเกิดเซลล์รับแสงแผ่นหนึ่ง[10][A]
กำเนิดเดียวหรือหลากหลาย[แก้]
การพิจารณาว่าตาได้วิวัฒนาการขึ้นครั้งหนึ่งหรือหรือหลายครั้ง จะขึ้นอยู่กับนิยามของตา กลไกทางพันธุกรรมที่ใช้ในพัฒนาการของตาจะเหมือนกันในสัตว์มีตาทั้งหมด ซึ่งอาจบ่งชี้ว่า บรรพบุรุษร่วมกันได้มีกลไกที่ไวแสงในบางรูปแบบ แม้จะยังไม่มีอวัยวะที่ใช้รับแสงโดยเฉพาะ
ถึงกระนั้น แม้แต่เซลล์รับแสงเองก็อาจจะวิวัฒนาการขึ้นมากกว่าครั้งเดียวจากตัวรับสารเคมีที่คล้ายคลึงกันทางโมเลกุล และเซลล์ไวแสงก็น่าจะมีก่อน เหตุการณ์ระเบิดสิ่งมีชีวิตยุคแคมเบรียนตั้งนาน[11] ความคล้ายคลึงกันในโครงสร้างระดับสูง เช่นการใช้โปรตีน crystallin เพื่อเป็นเลนส์ที่เกิดอย่างเป็นอิสระระหว่างเซฟาโลพอดและสัตว์มีกระดูกสันหลัง[12] สะท้อนให้เห็นกระบวนการ co-option ที่เปลี่ยนใช้โปรตีนซึ่งเคยมีบทบาทพื้นฐานอย่างหนึ่งเพื่อใช้สำหรับหน้าที่อีกอย่างหนึ่งในตา[13]
ลักษณะสืบสายพันธุ์ที่แชร์ร่วมกันระหว่างอวัยวะไวแสงทั้งหมด รวมกลุ่มโปรตีนไวแสงที่เรียกว่า อ็อปซิน (opsin) ซึ่งแบ่งออกเป็น 9 กลุ่ม และมีอยู่แล้วในเคลด urbilaterian ซึ่งเป็น บรรพบุรุษร่วมกันหลังสุดของสัตว์ที่ข้างซ้ายขวาสมมาตรกัน (ไบลาทีเรีย)[14] นอกจากนั้น ยังมีเครื่องมือทางพันธุกรรมเพื่อจัดตำแหน่งตาซึ่งสามัญในสัตว์ทั้งหมด คือ ยีน PAX6 ซึ่งควบคุมว่าตาจะพัฒนาขึ้นที่ตรงไหนในสัตว์ต่าง ๆ เริ่มตั้งแต่หมึก[15] จนถึงหนูหริ่งและแมลงวันทอง[16][17][18]
ดังนั้นโดยนัย ยีนสำหรับโครงสร้างระดับสูงเช่นนี้ จึงต้องเก่าแก่กว่าโครงสร้างที่พวกมันควบคุมดังที่เห็นในปัจจุบัน พวกมันดั้งเดิมจึงต้องมีหน้าที่ต่างกัน ก่อนจะนำมาใช้เพื่อบทบาทใหม่ในพัฒนาการของตา[13] อวัยวะรับความรู้สึกน่าจะวิวัฒนาการเกิดก่อนสมอง เพราะอวัยวะประมวลข้อมูล (คือสมอง) ย่อมไม่จำเป็นก่อนมีข้อมูลสำหรับแปลผล[19]
ระยะต่าง ๆ ของวิวัฒนาการตา[แก้]

องค์ประกอบแรกสุดของตา ก็คือโปรตีนไวแสง ซึ่งพบแม้แต่ในสิ่งมีชีวิตเซลล์เดียวโดยเรียกว่า จุดรับแสง (eyespot) จุดรับแสงจะสามารถรู้ว่ามีแสงรอบ ๆ คือสามารถแยกแยะความมืดจากสว่าง ซึ่งพอให้สัตว์ตอบสนองต่องช่วงแสง (photoperiodism) และต่อสนองต่อแสงแบบจังหวะเซอร์เคเดียน (circadian rhythm) แต่ก็ไม่พอต่อการเห็น เพราะไม่สามารถแยกแยะรูปร่างหรือทิศทางของแสง
จุดรับแสงจะพบในกลุ่มสัตว์ใหญ่ ๆ เกือบทั้งหมด และสามัญในบรรดาสิ่งมีชีวิตเซลล์เดียวรวมทั้งยูกลีนา จุดรับแสงของยูกลีนาเรียกอีกอย่างหนึ่งว่า "stigma" (แปลว่า จุดรับแสงเหมือนกัน) ซึ่งอยู่ที่ปลายเซลล์ด้านหน้า เป็นจุดรงควัตถุสีแดงซึ่งบังผลึกไวแสงจำนวนหนึ่ง เมื่อรวมกับแฟลเจลลัมด้านหน้า จุดไวแสงจะช่วยให้สิ่งมีชีวิตเคลื่อนไหวตอบสนองต่อแสง บ่อยครั้งไปทางแสงเพื่ออำนวยการสังเคราะห์แสง[20] และเพื่อกำหนดกลางวันกลางคืน โดยเป็นส่วนของจังหวะเซอร์เคเดียน
สารรงควัตถุสำหรับการเห็นจะอยู่ในสมองของสิ่งมีชีวิตที่ซับซ้อนกว่า และเชื่อว่า มีบทบาทในการประสานเวลาการวางไข่กับโคจรพระจันทร์ เพราะสามารถตรวจจับความเปลี่ยนแปลงเล็ก ๆ น้อย ๆ ของแสงในเวลากลางคืน สิ่งมีชีวิตจึงสามารถประสานเวลาปล่อยตัวอสุจิและไข่ เพื่อให้มีโอกาสผสมพันธุ์อย่างสูงสุด[ต้องการอ้างอิง]
ส่วนการเห็นเองจะอาศัยกลไกทางเคมี-ชีวภาพพื้นฐานที่สามัญต่อตาทุก ๆ แบบ แต่การสร้างโครงสร้างเพื่อตีความสิ่งแวดล้อมโดยมีกลไกนี้เป็นมูลฐานของสิ่งมีชีวิต จะต่างกันอย่างมาก คือตามีโครงสร้างและรูปแบบต่าง ๆ มากมาย โดยทั้งหมดเกิดวิวัฒนาการขึ้นค่อนข้างช้าเมื่อเทียบกับโปรตีนและโมเลกุลที่เป็นมูลฐาน[20]
ในระดับเซลล์ ดูเหมือนจะมีตาสองรูปแบบ สัตว์เคลด protostome รวมทั้งมอลลัสกา สัตว์พวกหนอนปล้อง และสัตว์ขาปล้อง จะมีรูปแบบหนึ่ง ส่วนสัตว์เคลด deuterostome รวมทั้งสัตว์มีแกนสันหลังและอิคีเนอเดอร์เมอเทอจะมีอีกรูปแบบหนึ่ง[20] หน่วยปฏิบัติการของตาก็คือเซลล์รับแสง ซึ่งมีโปรตีนอ็อปซินและตอบสนองต่อแสงโดยสร้างกระแสประสาท และอ็อปซินไวแสงจะอยู่ในชั้นเยื่อที่เป็นขนเพื่อให้มีพื้นที่ผิวมากที่สุด แต่ "ขน" ก็ยังมีลักษณะต่าง ๆ กันโดยรูปแบบหลักสองอย่างจะเป็นมูลฐานโครงสร้างของเซลล์รับแสง คือ microvilli และ cilia[21]
ตาของสัตว์เคลด protostome จะมี microvilli ซึ่งเป็นส่วนยื่นออกจากเยื่อหุ้มเซลล์ แต่ตาของสัตว์เคลด deuterostome จะมี cilia ซึ่งเป็นโครงสร้างต่างหากโดยไม่ใช่ส่วนของเยื่อหุ้มเซลล์[20] อนึ่ง ที่นอกตา สิ่งมีชีวิตอาจใช้เซลล์รับแสงแบบอื่น ๆ เช่น สัตว์พวกหนอนปล้องสปีชีส์ Platynereis dumerilii จะใช้ microvilli ในตา แม้จะมีเซลล์รับแสงแบบมี cilia ในสมองส่วนลึก[22]
ความเป็นอนุพันธ์ระหว่างโครงสร้างสองอย่างนี้อาจจะซับซ้อนคือ microvilli บางชนิดดูจะมีร่องรอยของ cilia แต่งานอื่น ๆ ก็ดูเหมือนจะสนับสนุนว่านี่เป็นความต่างกันแบบพื้นฐานระหว่างสัตว์เคลด protostome และ deuterostome[20] ข้อพิจารณาเหล่านี้มาจากการตอบสนองของเซลล์ต่อแสง โดยบางอย่างใช้โซเดียมสร้างสัญญาณไฟฟ้าเป็นกระแสประสาท และอย่างอื่นก็ใช้โพแทสเซียม นอกจากนั้น protostome โดยทั่วไปจะสร้างสัญญาณโดยปล่อยให้โซเดียมซึมผ่านผนังเซลล์ "มากกว่า" เทียบกับ deuterostome ที่ปล่อยน้อยกว่า[20] ซึ่งแสดงนัยว่า เมื่อสายพันธุ์ของสัตว์สองจำพวกนี้แยกออกจากกันก่อนยุคแคมเบรียน พวกมันมีเซลล์รับแสงที่ค่อนข้างพื้นฐานมาก แล้วจึงพัฒนามาเป็นตาที่ซับซ้อนยิ่งขึ้นโดยแยกกันต่างหาก
ตายุคต้น ๆ[แก้]
หน่วยประมวลแสงพื้นฐานของตาก็คือเซลล์รับแสง ซึ่งเป็นเซลล์พิเศษที่มีโมเลกุลสองอย่างในเยื่อหุ้มเซลล์ คือโปรตีนไวแสงอ็อปซิน ซึ่งล้อมรอบสารรงควัตถุ chromophore ที่ช่วยแยกแยะสี กลุ่มเซลล์เช่นนี้จะเรียกว่า จุดไวแสง (eyespots) ซึ่งวิวัฒนาการเกิดขึ้นเองกว่า 40-65 ครั้ง จุดไวแสงจะช่วยให้สัตว์รับรู้ทิศทางและความสว่างโดยพื้นฐาน แต่จะไม่พอแยกแยะวัตถุต่าง ๆ จากสิ่งแวดล้อม[20] การพัฒนาระบบที่สามารถแยกแยะทิศทางของแสงที่ละเอียดจนถึงไม่กี่องศาดูเหมือนจะเป็นเรื่องยากกว่ามาก และไฟลัมสัตว์เพียงแค่ 6 ไฟลัมจาก 30กว่า ไฟลัม[B] ได้พัฒนาระบบเช่นนี้ แต่สปีชีส์สิ่งมีชีวิต 96% ทั้งหมดในปัจจุบันก็อยู่ในไฟลัมเหล่านี้[20]

ระบบรับแสงที่ซับซ้อนเช่นนี้เริ่มต้นจากการเป็นแผ่นไวแสงมีหลายเซลล์ซึ่งค่อย ๆ ยุบลงเป็นรูปถ้วย ซึ่งแรกสุดจะทำให้สามารถแยกแยะทิศทางของแสงได้ดีขึ้น โดยจะรู้ทิศทางได้ละเอียดยิ่งขึ้น ๆ เมื่อหลุมลึกลงเรื่อย ๆ เทียบกับแผ่นไวแสงแบน ๆ ที่ไม่สามารถกำหนดทิศทางของแสง เพราะแสงไม่ว่าจะมาจากทิศทางไหนก็จะกระตุ้นเซลล์รับแสงเดียวกัน ตาที่มีรูปถ้วยจะช่วยให้แยกแยะทิศทางได้บ้าง เพราะแสงในทิศต่าง ๆ จะกระตุ้นเซลล์รับแสงที่ต่างกัน ตาเป็นหลุม ซึ่งเกิดขึ้นโดยยุคแคมเบรียน จะเห็นได้ในหอยทากดึกดำบรรพ์[โปรดขยายความ] และก็พบด้วยในหอยทากและสัตว์ไม่มีกระดูกสันหลังที่ยังมีอยู่ทุกวันนี้ เช่น พลานาเรีย
พลานาเรียสามารถแยะแยะทิศทางและระดับแสงได้เล็กน้อยเพราะมีเซลล์จอตาเป็นรูปถ้วยที่มีสารรงควัตถุอยู่อย่างหนาแน่น ซึ่งช่วยกันเซลล์ไวแสงจากแสงทุก ๆ ทิศทางยกเว้นจากช่องเดียวที่เปิดให้แสงเข้า อย่างไรก็ดี ตาต้นแบบเช่นนี้ก็ยังมีประสิทธิภาพในการตรวจจับความมีหรือไม่มีแสงได้ดีกว่าตรวจจับทิศทางของแสง โดยสถานการณ์นี้จะค่อย ๆ เปลี่ยนไปเมื่อหลุมตาลึกลงเรื่อย ๆ และมีจำนวนเซลล์รับแสงเพิ่มขึ้น ซึ่งทำให้ได้ข้อมูลทางตาที่แม่นยำเพิ่มขึ้น[23]
ในสัตว์ที่ซับซ้อนกว่านั้น เมื่อ chromophore ดูดกลืนโฟตอน ปฏิกิริยาเคมีจะถ่ายโอนพลังงานของโฟตอนเป็นพลังงานไฟฟ้า แล้วส่งต่อไปยังระบบประสาท เซลล์รับแสงจะเป็นส่วนของจอตา ซึ่งเป็นชั้นเซลล์บาง ๆ ที่ส่งข้อมูลทางตา[24] รวมทั้งข้อมูลแสงและข้อมูลเกี่ยวกับระยะช่วงวัน ซึ่งจำเป็นสำหรับระบบจังหวะเซอร์เคเดียนในสมอง อย่างไรก็ดี ก็ยังมีแมงกะพรุนบางชนิด เช่นในสกุล Cladonema ซึ่งมีตาที่ซับซ้อนแต่ไม่มีสมอง ตาจะส่งกระแสประสาทไปยังกล้ามเนื้อโดยตรงโดยไม่มีสมองที่ช่วยประมวลผลในระหว่าง[19]
ในช่วงเหตุการณ์ระเบิดสิ่งมีชีวิตยุคแคมเบรียน การพัฒนาของตาได้เร่งขึ้นอย่างรวดเร็ว โดยมีการประมวลภาพและการตรวจจับทิศทางของแสงเป็นการเปลี่ยนแปลงใหญ่ ๆ[25]
.jpg)
หลังจากเขตเซลล์ไวแสงกลายเป็นรูปถ้วย ก็จะมาถึงจุดที่การลดขนาดของรูแสง จะมีประสิทธิภาพเพิ่มรายละเอียดของภาพมากกว่าที่ถ้วยรับแสงจะลึกลงเรื่อย ๆ[10] โดยลดขนาดรูแสง สัตว์ก็จะสามารถสร้างภาพได้จริง ๆ ช่วยให้รับรู้ทิศทางแสงได้ละเอียดขึ้น พร้อมกับการรับรู้รูปร่างโดยบางส่วน ตาชนิดนี้พบในหอยงวงช้าง เป็นตาที่ไม่มีกระจกตาหรือแก้วตา (เลนส์ตา) ซึ่งทำให้มีรายละเอียดน้อยและเห็นภาพมัว ๆ แต่ก็ยังเห็นดีกว่าแผ่นไวแสงในยุคต้น ๆ[26] ต่อมาการงอกเกินของเซลล์โปร่งแสงจึงช่วยกันการเปื้อนและปรสิต
เมื่อช่องต่าง ๆ ของตาเกิดแยกจากกัน ก็จะค่อย ๆ เปลี่ยนเกิดสารน้ำพิเศษในลูกตา เพื่อให้เกิดความเหมาะสมสูงสุดในการกรองแสง ในการกันแสงอัลตราไวโอเลต ในการทำงานในน้ำหรือนอกน้ำ และเพื่อดรรชนีหักเหแสงที่เหมาะสุด ในสัตว์บางชั้น ชั้นต่าง ๆ ของตาอาจสัมพันธ์กับคราบหรือเปลือกนอกของสัตว์ ตัวอย่างนี้เห็นได้ในหนอนกำมะหยี่ที่เปลือกนอก (cuticula) จะเข้าเป็นส่วนของกระจกตาด้วย คือกระจกตาจะประกอบด้วยชั้น cuticula เดียวหรือสองชั้นขึ้นอยู่กับว่า หนอนได้ลอกคราบเร็ว ๆ นี้หรือไม่[27] พร้อมกับเลนส์ตาและสารน้ำในลูกตาสองส่วน กระจกตาจะมีหน้าที่รวมแสง ช่วยโฟกัสแสงที่จอตา และป้องกันลูกตา โดยให้กำลังหักเหแสงเป็น 2/3 ของตาทั้งหมด[28]
เหตุผลหลักที่ตามีหน้าที่เฉพาะเพื่อตรวจจับสเปกตรัมแม่เหล็กไฟฟ้าที่มีความยาวคลื่นในพิสัยจำกัด (คือ สเปกตรัมที่มองเห็นได้) อาจเป็นเพราะสปีชีส์แรก ๆ สุดที่ไวแสงเป็นสัตว์น้ำ และคลื่นแม่เหล็กไฟฟ้าที่มีความยาวคลื่นโดยเฉพาะสองชนิด คือ สีน้ำเงินและสีเขียวที่มองเห็นได้ สามารถผ่านเข้าไปในน้ำได้ โดยคุณสมบัติกรองแสงของน้ำเช่นนี้ ก็มีอิทธิพลต่อความไวแสงของพืชต่าง ๆ ด้วย[29][30][31]
วิวัฒนาการของเลนส์และความหลากหลาย[แก้]

ในตาที่ไร้เลนส์ แสงจากจุดไกลจะตกลงที่หลังตาโดยมีขนาดประมาณเท่ากับรูรับแสงของตา เมื่อมีเลนส์ แสงก็จะตกลงที่พื้นผิวซึ่งเล็กกว่า โดยไม่ได้ลดระดับความชัดเจนของสิ่งเร้า[6] ความยาวโฟกัสของสัตว์คล้ายหนอนกลุ่ม lobopodia ที่มีตาง่าย ๆ พร้อมกับเลนส์ จะโฟกัสภาพที่ "หลัง" จอตา ดังนั้น แม้จะไม่สามารถมองเห็นภาพได้ชัด แต่ความเข้มของแสงจะทำให้สัตว์สามารถเห็นได้ไกลขึ้นและมองเห็นในน้ำที่มืดได้ดีขึ้น[27] การเพิ่มดรรชนีหักเหแสงของเลนส์ต่อมา น่าจะมีผลเป็นการมีภาพที่ได้โฟกัส[27]
ส่วนการพัฒนาเลนส์ตาในรูปแบบคล้ายกับกล้องถ่ายรูป น่าจะมาจากอีกเส้นทางหนึ่ง เซลล์โปร่งแสงที่ปกคลุมช่องรูเข็มของตา ได้แบ่งออกเป็นสองชั้นโดยมีน้ำอยู่ในระหว่าง[ต้องการอ้างอิง] ซึ่งตอนแรกไหลเวียนถ่ายออกซิเจน สารอาหาร ของเสีย และมีหน้าที่ทางภูมิคุ้มกัน ทำให้เลนส์หนาขึ้นและสามารถป้องกันตาได้ดีขึ้น ยิ่งกว่านั้น การต่อประสานระหว่างของเแข็งของเหลวเป็นหลาย ๆ ชั้น ก็จะเพิ่มกำลังหักเหแสงของเลนส์ ทำให้มีมุมมองกว้างขึ้นและมีภาพชัดขึ้น และเช่นกันอีก การแบ่งออกเป็นชั้น ๆ ของเลนส์ก็อาจเกี่ยวกับการลอกคราบ น้ำในเซลล์อาจจะไหลเต็มระหว่างชั้นเองตามธรรมชาติ[ต้องการอ้างอิง] ให้สังเกตว่า โครงสร้างทางตาเช่นนี้โดยเฉพาะยังไม่เคยพบในสิ่งมีชีวิต และก็ไม่คิดว่าจะพบในอนาคต นอกจากนั้น ซากดึกดำบรรพ์แทบไม่เคยรักษาเนื้อเยื่ออ่อน ๆ ไว้ได้ และแม้ว่าได้ ช่องของเหลวใหม่ก็น่าจะปิดไปเมื่อซากแห้ง หรือเมื่อตะกอนทับถมกดทับให้ชั้นต่าง ๆ รวมตัวกัน ทำให้ตาดูเหมือนกับโครงสร้างที่มีในรุ่นก่อน

แก้วตา (เลนส์ตา) ของสัตว์เลี้ยงลูกด้วยนม จะประกอบด้วยเซลล์เยื่อบุผิวพิเศษที่มีโปรตีน crystallin อยู่อย่างหนาแน่น โดยมีโปรตีนในสองกลุ่มหลัก ๆ คือ α-crystallin และ βγ-crystallin แม้จะเป็นโปรตีนที่ดั้งเดิมใช้ในหน้าที่อื่น ๆ แต่ในที่สุดก็ปรับใช้เพื่อการเห็นโดยตาของสัตว์เพียงเท่านั้น[32]
ในตัวอ่อน แก้วตาจะเป็นเนื้อเยื่อที่ยังมีชีวิต แต่ส่วนต่าง ๆ ของเซลล์ก็ยังไม่โปร่งแสง และดังนั้น จึงต้องเอาออกก่อนที่สัตว์จะมองเห็นได้ เมื่อเอาออกแล้ว นี่ก็จะเป็นเซลล์ที่ตายแล้วและเต็มไปด้วย crystallin โปรตีนนี้พิเศษ โดยมีคุณสมบัติโดยเฉพาะที่จำเป็นเพื่อความโปร่งแสงและการทำหน้าที่เป็นแก้วตา เช่น อัดแน่น ทนทานต่อการแปรเปลี่ยนเป็นผลึก และอายุยืนนาน เพราะมันจะต้องใช้ได้ตลอดชีวิตของสัตว์[32] เกรเดียนต์ดรรชนีหักเหแสงซึ่งทำให้เลนส์มีประโยชน์ เกิดจากความเปลี่ยนแปลงแบบเป็นรัศมี (radial) ของความหนาแน่นของ crystallin ในส่วนต่าง ๆ ของเลนส์ ไม่ใช่เพราะมีโปรตีนชนิดใดชนิดหนึ่งโดยเฉพาะ คือ มันไม่ใช่เนื่องจากการมี crystallin แต่เป็นการกระจายตัวของมันโดยเปรียบเทียบในส่วนต่าง ๆ ของเลนส์ ที่ทำให้เลนส์ใช้งานได้[33]
มันเป็นเรื่องยากทางชีวภาพเพื่อดำรงชั้นเซลล์โปร่งแสง ดังนั้น การพอกสะสมวัสดุโปร่งแสงที่ไม่มีชีวิต ทำให้ไม่จำเป็นต้องส่งสารอาหารหรือกำจัดของเสีย สัตว์ทะเลโบราณไทรโลไบต์ใช้แคลไซต์ ซึ่งเป็นแร่ที่ปัจจุบันใช้เพื่อมองเห็นเพียงในสปีชีส์เดียวของดาวเปราะ[34] ในตาประกอบอื่น ๆ [ต้องการตรวจสอบความถูกต้อง] และตาที่คล้ายกล้องถ่ายรูป วัสดุที่จะใช้ก็คือ crystallin ช่องในระหว่างชั้นเนื้อเยื่อตามธรรมชาติ จะทำให้เลนส์มีรูปนูนออกสองข้าง ซึ่งเป็นรูปร่างอุดมคติทางแสงและทางกลศาสตร์สำหรับวัสดุที่มีดรรชนีหักเหแสงธรรมดา[โปรดขยายความ] เลนส์ที่นูนออกทั้งสองข้างไม่เพียงอำนวยภาพที่ละเอียด แต่ยังอำนวยรูรับแสงและภาพในแสงสลัว เพราะความละเอียดของภาพจะไม่จับคู่กับขนาดรูรับแสงอีกต่อไป รูรับแสงจึงเริ่มขยายขนาดขึ้นอีกอย่างช้า ๆ เพราะมีข้อจำกัดทางการไหลเวียนที่ลดลง

อย่างเป็นอิสระเอง ชั้นโปร่งแสงและชั้นทึบอาจแยกออกมาจากเลนส์ทางด้านหน้า คือ กระจกตาและม่านตา (iris) (ซึ่งอาจเกิดขึ้นก่อนหรือหลังการพอกสะสมผลึกโปร่งแสง) การแยกชั้นออกทางด้านหน้าอีกต่อมาก็สร้างสารน้ำในลูกตา (aqueous humour) ซึ่งเพิ่มดรรชนีหักเหแสงและลดปัญหาการไหลเวียนลงอีก
การเกิดเนื้อเยื่อที่ไม่โปร่งแสงรอบ ๆ เป็นรูปแหวน ก็ทำให้มีหลอดเลือดได้มากขึ้น มีการไหลเวียนมากขึ้น และทำให้ตาขยายขนาดได้อีก และแผ่นเนื้อเยื่อรอบ ๆ เลนส์เช่นนี้ยังช่วยอำพรางข้อบกพร่องของเลนส์ที่สามัญตรงส่วนรอบ ๆ อีกด้วย ความจำเป็นเพื่ออำพรางข้อบกพร่องของเลนส์ก็เพิ่มขึ้นเรื่อย ๆ ตามเลนส์ที่โค้งนูนและมีกำลังเพิ่มขึ้น ตามขนาดเลนส์และตาที่ใหญ่เพิ่มขึ้น เพื่อให้มีรายละเอียดภาพและรูรับแสงที่เพิ่มขึ้นของสัตว์ โดยทั้งหมดมีแรงกดดันจากความจำเป็นในการล่าหรือการรอดชีวิต ระบบตาเช่นนี้ปัจจุบันมีลักษณะหน้าที่คล้าย ๆ กันในสัตว์มีกระดูกสันหลังทั้งหมดรวมทั้งมนุษย์ด้วย นักวิชาการได้สรุปไวว้า "รูปแบบพื้นฐานของตาสัตว์มีกระดูกสันหลังทั้งหมดคล้ายกัน"[35]
พัฒนาการอื่น ๆ[แก้]
มีสารรงควัตถุ 5 กลุ่มที่พบในสัตว์มีกระดูกสันหลัง ทั้ง 5 กลุ่มยกเว้นประเภทเดียวได้วิวัฒนาการเกิดขึ้นก่อนการแยกสายพันธุ์ระหว่าง cyclometer และปลา[36]
การปรับใช้สารรงควัตถุ 5 กลุ่มนี้ทำให้เกิดตาที่เหมาะสมกับสเปกตรัมแม่เหล็กไฟฟ้าที่สัตว์ประสบ คือ เมื่อแสงวิ่งผ่าน น้ำจะดูดซึมคลื่นแสงที่ยาวกว่า เช่นสีแดงและเหลือง ได้เร็วกว่าคลื่นที่สั้นกว่า เช่นสีเขียวและสีน้ำเงิน ดังนั้น เมื่อน้ำลึกขึ้น ก็จะทำให้เกิดชั้นแสงต่าง ๆ ตัวรับแสงของปลาจะไวต่อพิสัยแสงที่พบในที่ที่ตนอยู่มากกว่า
แต่ปรากฏการณ์เช่นนี้ไม่มีบนบก ทำให้สัตว์มีกระดูกสันหลังบนบกไม่ค่อยไวแสงในพิสัยต่าง ๆ กัน การเห็นสีได้คล้าย ๆ กันจึงมีผลให้เกิดการสื่อสารผ่านสีอย่างสำคัญ[36] ซึ่งทำให้สัตว์ที่เห็นสีต่าง ๆ ได้เปรียบทางการคัดเลือกมากกว่า เช่น รู้จักสัตว์ล่าเหยื่อ อาหาร และคู่ได้ดีกว่า นักวิชาการบางพวกเชื่อว่า กลไกทางการรับรู้-ประสาทจะเป็นตัวควบคุมรูปแบบทางพฤติกรรมทั่วไป เช่น การหนี การหาอาหาร และการซ่อนตัว รูปแบบพฤติกรรมที่เฉพาะเจาะจงต่อพิสัยความยาวแสงได้พบและแบ่งเป็น 2 กลุ่มหลัก ๆ คือ ที่ความยาวคลื่นน้อยกว่า 450 นาโนเมตร จะสัมพันธ์กับแสงธรรมชาติ และที่ความยาวคลื่นมากกว่า 450 นาโนเมตร จะสัมพันธ์กับแสงสะท้อน[37]
เพราะโมเลกุลอ็อปซินสามารถตรวจจับความยาวคลื่นแสงที่ต่างกันเล็ก ๆ น้อย ๆ ในที่สุด การเห็นเป็นสีก็เกิดขึ้น เมื่อเซลล์รับแสงมีอ็อปซินไวแสงที่มีความยาวคลื่นต่าง ๆ กัน[24] เนื่องจากเป็นการปรับตัวทางเคมี ไม่ใช่ทางกลศาสตร์ นี่อาจจะเกิดขึ้นในระยะต้น ๆ ของวิวัฒนาการทางตา และสมรรถภาพนี้ก็อาจจะหายไปแล้วเกิดขึ้นอีกเมื่อสัตว์กลายเป็นสัตว์ล่าหรือสัตว์เหยื่อ และเช่นกัน การเห็นเวลากลางวันและกลางคืนก็เกิดขึ้นเมื่อเซลลรับแสงแยกออกเป็นเซลล์รูปกรวยและเซลล์รูปแท่ง[ต้องการอ้างอิง]
การเห็นแสงโพลาไรซ์[แก้]
ดังที่กล่าาวแล้ว คุณสมบัติของแสงใต้น้ำต่างจากในอากาศ ตัวอย่างหนึ่งก็คือปรากฏการณ์โพลาไรเซชันของแสง ซึ่งเป็นการจัดระเบียบแสงจากพระอาทิตย์ในเชิงเส้น (แบบลินเนียร์) และเกิดเมื่อแสงวิ่งผ่านช่องเล็กยาวเช่นฟิลเตอร์ หรือเมื่อวิ่งผ่านเข้าตัวกลาง/สื่ออีกชนิดหนึ่ง
ความไวแสงโพลาไรซ์มีประโยชน์เป็นพิเศษต่อสิ่งมีชีวิตที่อยู่ใต้น้ำซึ่งลึกกว่า 2-3 เมตร เพราะในสิ่งแวดล้อมเช่นนี้ การเห็นเป็นสีพึ่งพาได้น้อยกว่า และดังนั้น จึงสร้างความกดดันทางการคัดเลือกเพื่อเห็นเป็นสีที่อ่อนกว่า แม้เซลล์รับแสงโดยมากจะมีสมรรถภาพในการแยกแยะแสงโพลาไรซ์ได้บ้าง แต่เซลล์ของสัตว์มีกระดูกสันหลังบนบกจะมีผิวเยื่อหุ้มเซลล์จัดในแนวตั้งฉากกับแสงที่วิ่งมากระทบ โดยมีลำดับแถวของส่วนกำเนิดสี (chromophore) ตามแกนต่าง ๆ โดยสุ่ม ซึ่งมีผลให้สัตว์ไม่ไวแสงโพลาไรซ์[38] แต่ปลาและนกบางชนิดก็ยังสามารถแยกแยะแสงโพลาไรซ์ได้ แสดงว่ามีเซลล์รับแสงที่มีบทบาทในการวิเคราะห์แสงโพลาไรซ์ในเชิงเส้น นอกจากนั้น หมึกกระดองก็สามารถเห็นแสงโพลาไรซ์ได้อย่างชัดเจน แม้จะไม่เห็นเป็นสี[39]
เหมือนกับการเห็นเป็นสี ความไวแสงโพลาไรซ์สามารถช่วยสิ่งมีชีวิตให้แยกแยะวัตถุและสัตว์อื่นที่อยู่รอบ ๆ ตัว เพราะแสงโพลาไรซ์สร้างแสงสะท้อนรบกวนน้อยมาก จึงสามารถใช้ในการกำหนดทิศทาง การนำทาง และการหาเหยื่อที่ซ่อนอยู่[38]
กลไกการโฟกัสแสง[แก้]
โดยใช้กล้ามเนื้อหูรูดของม่านตา (iris sphincter muscle) สัตว์บางชนิดจะสามารถขยับแก้วตาไปทางด้านหน้าหลังได้ และสัตว์บางชนิดจะดึงเลนส์ให้แบนลง การควบคุมขนาดของตาและดำรงความยาวโฟกัสเป็นกลไกทางเคมีอีกอย่างต่างหาก รูปร่างของรูม่านตา (pupil) จะสามารถใช้พยากรณ์ระบบโฟกัสที่ตาใช้ รูม่านตาแบบช่องแคบยาวแสดงระบบควบคุมโฟกัสจากหลายจุด แบบกลมปกติจะแสดงระบบควบคุมโฟกัสจากจุดเดียว ในแบบกลม รูม่านตาจะลดขนาดในแสงสว่างแล้วเพิ่มความยาวโฟกัส และจะขยายขนาดในที่มืดแล้วลดความยาวโฟกัส[40]
ให้สังเกตว่า การโฟกัสได้ไม่ใช่เรื่องคอขาดบาดตาย ดังที่ช่างกล้องจะรู้ ความผิดพลาดในการโฟกัสจะเพิ่มเมื่อรูรับแสงใหญ่ขึ้น ดังนั้น มีสัตว์จำนวนนับไม่ได้ที่มีตาเล็ก แต่หากินในเวลากลางวันและสามารถรอดชีวิตโดยไม่มีกลไกโฟกัสตาเลย เมื่อสัตว์มีขนาดใหญ่ขึ้น หรือเปลี่ยนไปอยู่ในที่มืดลง ความจำเป็นในการโฟกัสแสงได้ก็จะเริ่มมีขึ้น
ตำแหน่ง[แก้]
สัตว์ที่เป็นเหยื่อทั่วไปจะมีตาทางด้านข้างศีรษะ เพื่อให้มีมุมการเห็นที่กว้างกว่าและช่วยให้หนีสัตว์ล่าเหยื่อได้ แต่สัตว์ล่าเหยื่อจะมีตาด้านหน้าเพื่อให้รู้ใกล้ไกลได้ดีกว่า[41][42]
ปลาซีกเดียวเป็นสัตว์ล่าเหยื่อซึ่งนอนตะแคงที่ก้นแหล่งน้ำ และมีตาทั้งสองข้างอยู่ทางด้านเดียวกันของหัว แต่ซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพของปลาจากสัตว์ที่มีสองข้างเท่ากันมากกว่าก็คือ Amphistium ซึ่งมีตาข้างหนึ่งที่กลางหัว
ของเหลือทางวิวัฒนาการ (Evolutionary baggage)[แก้]

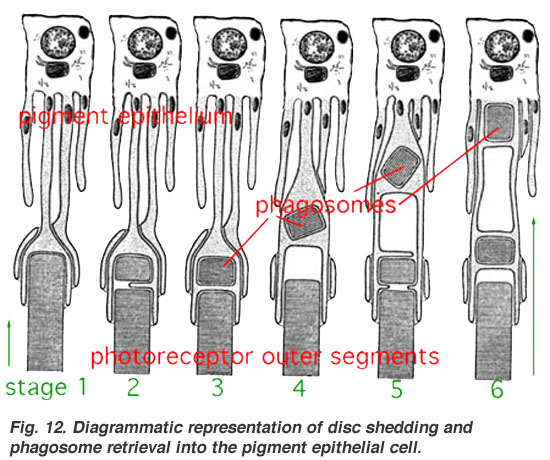
ตาของสัตว์จำนวนมากมีกายวิภาคซึ่งเหมือนกับบันทึกทางประวัติวิวัฒนาการของตาด้วย ยกตัวอย่างเช่น ตาของสัตว์มีกระดูกสันหลัง สร้างขึ้น "กลับหน้ากลับหลังและตีลังกา" ซึ่งบังคับให้ "โฟตอนแสงต้องวิ่งผ่านกระจกตา, แก้วตา, น้ำ, เส้นเลือด, เซลล์ปมประสาท (ganglion cell), amacrine cell, horizontal cell, bipolar cell ก่อนจะไปถึงเซลล์รูปแท่งและเซลล์รูปกรวยที่ไวแสงและถ่ายโอนสัญญาณแสงให้เป็นกระแสประสาท แล้วส่งต่อไปยังเปลือกสมองส่วนการเห็นที่สมองด้านหลังเพื่อประมวลหารูปแบบต่าง ๆ ที่สำคัญต่อสิ่งมีชีวิต"[43] แม้โครงสร้างเช่นนี้จะมีจุดอ่อนบ้าง แต่ก็อำนวยผิวจอตาของสัตว์มีกระดูกสันหลังให้สามารถดำรงเมแทบอลิซึมระดับสูง เทียบกับโครงสร้างที่ไม่กลับหน้ากลับหลัง[44] และอำนวยให้เกิดวิวัฒนาการของชั้นคอรอยด์ (Choroid) รวมทั้ง retinal pigment epithelial (RPE) cell ซึ่งมีบทบาทสำคัญในการป้องกันเซลล์รับแสงจากความเสียหายเพราะออกซิเดชันที่เนื่องกับแสง[45][46]
โดยเทียบกับแล้ว ตาของเซฟาโลพอด ได้เกิดแบบสมหน้าสมหลัง คือมีใยประสาทยื่นออกจากจอตาด้านหลัง ทำให้สัตว์ไม่มีจุดบอด ความแตกต่างนี้อาจมาจากกำเนิดตา เพราะในเซฟาโลพอด ตาเกิดจากการม้วนเข้าของผิวศีรษะ (head surface) เทียบกับสัตว์มีกระดูกสันหลัง ที่ตามาจากส่วนของสมอง[47]
เชิงอรรถ[แก้]
- ↑ David Berlinski, an intelligent design proponent, questioned the basis of the calculations, and the author of the original paper refuted Berlinski's criticism.
- Berlinski, David (April 2001). "Commentary magazine".
{{cite journal}}: Cite journal ต้องการ|journal=(help) - Nilsson, Dan-E. "Beware of Pseudo-science: a response to David Berlinski's attack on my calculation of how long it takes for an eye to evolve".
{{cite journal}}: Cite journal ต้องการ|journal=(help) - "Evolution of the Eye" on PBS
- Berlinski, David (April 2001). "Commentary magazine".
- ↑ ตัวเลขจะต่าง ๆ กันตามนักวิชาการ
อ้างอิง[แก้]
- ↑ Land, MF; Nilsson, D-E (2002). Animal Eyes. Oxford: Oxford University Press.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Lee, M. S. Y.; Jago, J. B.; Garcia-Bellido, D. C.; Edgecombe, G. E.; Gehling, J. G; Paterson, J. R. (2011). "Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia". Nature. 474: 631–634. doi:10.1038/nature10097. PMID 21720369.
- ↑ 3.0 3.1 Darwin, Charles (1859). On the Origin of Species. London: John Murray.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Gehring, WJ (2005). "New perspectives on eye development and the evolution of eyes and photoreceptors". J. Hered. 96 (3): 171–84. doi:10.1093/jhered/esi027. PMID 15653558.
- ↑ Nilsson, D.-E. (2013). "Eye evolution and its functional basis". Visual Neuroscience. 30: 5–20. doi:10.1017/s0952523813000035.
- ↑ 6.0 6.1 Nilsson, D.-E.; Pelger, S. (1994). "A pessimistic estimate of the time required for an eye to evolve". Proceedings of the Royal Society B: Biological Sciences. 256: 53–58. doi:10.1098/rspb.1994.0048. PMID 8008757.
- ↑ Young, G. C. (2008). "Early evolution of the vertebrate eye - fossil evidence". Evo Edu Outreach. 1: 427–438. doi:10.1007/s12052-008-0087-y.
- ↑ Parker, A. R. (2009). "On the origin of optics". Optics & Laser Technology. 43 (2): 323–329. Bibcode:2011OptLT..43..323P. doi:10.1016/j.optlastec.2008.12.020.
- ↑ Parker, Andrew (2003). In the Blink of an Eye: How Vision Sparked the Big Bang of Evolution. Cambridge, MA: Perseus Pub. ISBN 0-7382-0607-5.
- ↑ 10.0 10.1 Nilsson, D-E; Pelger S (1994). "A pessimistic estimate of the time required for an eye to evolve". Proceedings of the Royal Society B. 256 (1345): 53–58. Bibcode:1994RSPSB.256...53N. doi:10.1098/rspb.1994.0048. PMID 8008757. S2CID 13061351.
- ↑ Nilsson, D. E. (1996). "Eye ancestry: old genes for new eyes". Current Biology. 6 (1): 39–42. doi:10.1016/S0960-9822(02)00417-7. PMID 8805210.
- ↑ Zinovieva, R.; Piatigorsky, J.; Tomarev, S. I. (1999). "O-Crystallin, arginine kinase and ferritin from the octopus lens". Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology. 1431 (2): 512–517. doi:10.1016/S0167-4838(99)00066-7.
- ↑ 13.0 13.1 Scotland, R. W. (2010). "Deep homology: A view from systematics". BioEssays. 32 (5): 438–449. doi:10.1002/bies.200900175. PMID 20394064.
- ↑ Ramirez, MD; Pairett, AN; Pankey, MS; Serb, JM; Speiser, DI; Swafford, AJ; Oakley, TH (2016-10-26). "The last common ancestor of most bilaterian animals possessed at least 9 opsins". Genome Biology and Evolution: evw248. doi:10.1093/gbe/evw248.
- ↑ Yoshida, Masa-aki; Yura, Kei; Ogura, Atsushi (2014-03-05). "Cephalopod eye evolution was modulated by the acquisition of Pax-6 splicing variants". Scientific Reports. nature.com. 4. Bibcode:2014NatSR...4E4256Y. doi:10.1038/srep04256. PMC 3942700. PMID 24594543. สืบค้นเมื่อ 2014-06-30.
- ↑ Halder, G.; Callaerts, P.; Gehring, W. J. (1995). "New perspectives on eye evolution". Current Opinion in Genetics & Development. 5 (5): 602–609. doi:10.1016/0959-437X(95)80029-8. PMID 8664548.
- ↑ Halder, G.; Callaerts, P.; Gehring, W. (1995). "Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila". Science. 267 (5205): 1788–92. Bibcode:1995Sci...267.1788H. doi:10.1126/science.7892602. PMID 7892602.
- ↑ Tomarev, S. I.; Callaerts, P.; Kos, L.; Zinovieva, R.; Halder, G.; Gehring, W.; Piatigorsky, J. (1997). "Squid Pax-6 and eye development". Proceedings of the National Academy of Sciences of the United States of America. 94 (6): 2421–2426. Bibcode:1997PNAS...94.2421T. doi:10.1073/pnas.94.6.2421. PMC 20103. PMID 9122210.
- ↑ 19.0 19.1 Gehring, W. J. (2005-01-13). "New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors" (Full text). Journal of Heredity. Oxford Journals. 96 (3): 171–184. doi:10.1093/jhered/esi027. PMID 15653558. สืบค้นเมื่อ 2008-04-26.
- ↑ 20.0 20.1 20.2 20.3 20.4 20.5 20.6 20.7 Land, MF; Fernald, RD (1992). "The Evolution of Eyes". Annual Review of Neuroscience. 15: 1–29. doi:10.1146/annurev.ne.15.030192.000245. PMID 1575438.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Autrum, H (1979). "Introduction". ใน Autrum, H (บ.ก.). Comparative Physiology and Evolution of Vision in Invertebrates- A: Invertebrate Photoreceptors. Handbook of Sensory Physiology. Vol. VII/6A. New York: Springer-Verlag. pp. 6–9. ISBN 3-540-08837-7.
- ↑ Arendt, D.; Tessmar-Raible, K.; Snyman, H.; Dorresteijn, A.W.; Wittbrodt, J. (2004-10-29). "Ciliary Photoreceptors with a Vertebrate-Type Opsin in an Invertebrate Brain". Science. 306 (5697): 869–871. doi:10.1126/science.1099955. PMID 15514158.
- ↑ Eye-Evolution? เก็บถาวร 2012-09-15 ที่ เวย์แบ็กแมชชีน
- ↑ 24.0 24.1 Fernald, Russell D. (2001). The Evolution of Eyes: How Do Eyes Capture Photons? เก็บถาวร 2006-03-19 ที่ เวย์แบ็กแมชชีน Karger Gazette 64: "The Eye in Focus".
- ↑ Conway-Morris, S (1998). The Crucible of Creation. Oxford: Oxford University Press.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ ริชาร์ด ดอว์กินส์ (1986). The Blind Watchmaker. United States: Norton & Company. ISBN 0-393-31570-3.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ 27.0 27.1 27.2 Schoenemann, B.; Liu, J. N.; Shu, D. G.; Han, J.; Zhang, Z. F. (2008). "A miniscule optimized visual system in the Lower Cambrian". Lethaia. 42 (3): 265–273. doi:10.1111/j.1502-3931.2008.00138.x.
- ↑ Ali, MA; Klyne, MA (1985). Vision in vertebrates. New York: Plenum Press.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Fernald, Russell D. (2001). The Evolution of Eyes: Why Do We See What We See? เก็บถาวร 2006-03-19 ที่ เวย์แบ็กแมชชีน Karger Gazette 64: "The Eye in Focus".
- ↑
Fernald, Russell D (1998). Aquatic Adaptations in Fish Eyes. New York: Springer.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Fernald, RD (1997). "The evolution of eyes". Brain Behav. Evol. 50 (4): 253–9. doi:10.1159/000113339. PMID 9310200.
- ↑ 32.0 32.1 Slingsby, C.; Wistow, G. J.; Clark, A. R. (2013). "Evolution of crystallins for a role in the vertebrate eye lens". Protein Science. 22: 367–380. doi:10.1002/pro.2229.
- ↑ Fernald, Russell D. (2001). The Evolution of Eyes: Where Do Lenses Come From? Karger Gazette 64: "The Eye in Focus".
- ↑ Burgess, Daniel S (2001). "Brittle Star Features Calcite Lenses". Photonics Spectra.
{{cite web}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Ali, Mohamed Ather; Klyne, M. A. (1985). Vision in Vertebrates. New York: Plenum Press. p. 1. ISBN 0-306-42065-1.
- ↑ 36.0 36.1 Osorio, D; Vorobyev, M (2005). "Photo-receptor spectral sensitivities in terrestrial animals: adaptations for luminescence and color vision". Proc. R. Soc. B.
- ↑ Menzel, Randolf (1979). "Spectral Sensitivity and Color Vision in Invertebrates". ใน Autrum, H (บ.ก.). Comparative Physiology and Evolution of Vision in Invertebrates- A: Invertebrate Photo-receptors. Handbook of Sensory Physiology. Vol. VII/6A. New York: Springer-Verlag. pp. 504–506, 551–558. ISBN 3-540-08837-7.
- ↑ 38.0 38.1 Cronin, T; Shashar, N; Caldwell, R; Marshall, J; Cheroske, A; Chiou, T (2003). "Polarization vision and its role in biological signaling". Integr. Comp. Biol. 1.4 MBPDF
- ↑ Mäthger, Lydia M.; Barbosa, Alexandra; Miner, Simon; Hanlon, Roger T. (2006-05-01). "Color blindness and contrast perception in cuttlefish (Sepia officinalis) determined by a visual sensorimotor assay". Vision Research. 46 (11): 1746–1753. doi:10.1016/j.visres.2005.09.035. PMID 16376404.
- ↑ Malstrom, T; Kroger, R (2006). "pupil shape and lens optics in the eyes of terrestrial vertebrates". The Journal of Experimental Biology.
- ↑ "Carnivores". U.S. Department of the Interior, Bureau of Land Management. 14 December 2009. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 14 June 2011. สืบค้นเมื่อ 28 March 2011.
- ↑ Boroditsky, Lera (24 June 1999). "Light & Eyes: Lecture Notes". Lecture Notes. Stanford. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 5 July 2010. สืบค้นเมื่อ 11 May 2010.
- ↑ Hitchens, Christopher (2007). God is Not Great. p. 82.
'backwards and upside down', 'photons of light to travel through the cornea, lens, aqueous fluid, blood vessels, ganglion cells, amacrine cells, horizontal cells, and bipolar cells before they reach the light-sensitive rods and cones that transduce the light signal into neural impulses, which are then sent to the visual cortex at the back of the brain for processing into meaningful patterns.'
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Reichenbach, A; Bringmann, A (2010). Müller cells in the healthy and diseased retina. New York: Springer. pp. 15–20.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ "LIGHT-INDUCED DAMAGE to the RETINA". 2009-03-03. เก็บจากแหล่งเดิมเมื่อ 2017-10-18. สืบค้นเมื่อ 2018-02-17.
- ↑ Diagrammatic representation of disc shedding and phagosome retrieval into the pigment epithelial cell (JPG). WebVision, Medical school @ University of Utah.
- ↑ Serb, Jeanne M.; Eernisse, Douglas J. (2008-09-25). "Charting Evolution's Trajectory: Using Molluscan Eye Diversity to Understand Parallel and Convergent Evolution". Evolution: Education and Outreach. 1 (4): 439–447. doi:10.1007/s12052-008-0084-1. ISSN 1936-6426.
{kind=link}
อ่านเพิ่ม[แก้]
- Lamb TD, Collin SP, Pugh EN (December 2007). "Evolution of the vertebrate eye: opsins, photoreceptors, retina and eye cup". Nat. Rev. Neurosci. 8 (12): 960–76. doi:10.1038/nrn2283. PMC 3143066. PMID 18026166. Illustration. Review
- Lamb, TD (2011). "Evolution of the Eye" (PDF). Scientific American. 305 (1): 64–69. Bibcode:2011SciAm.305f..64L. doi:10.1038/scientificamerican0711-64. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 12 ธันวาคม 2013. สืบค้นเมื่อ 28 เมษายน 2013.
- Land, Michael F.; Nilsson, Dan-Eric (2012). "The origin of vision". Animal Eyes (2 ed.). Oxford: Oxford University Press. pp. 1–22. ISBN 978-0199581146.
- Journal Evolution: Education and Outreach[ลิงก์เสีย] Volume 1, Number 4 / October 2008. Special Issue: The Evolution of Eyes. 26 articles, free access.
- Ivan R. Schwab (2012). Evolution's Witness: How Eyes Evolved. New York: Oxford University Press. ISBN 9780195369748.
- Hayakawa S, Takaku Y, Hwang JS, Horiguchi T, Suga H, Gehring W, และคณะ (2015). "Function and evolutionary origin of unicellular camera-type eye structure". PLOS ONE. 10 (3): e0118415. Bibcode:2015PLoSO..1018415H. doi:10.1371/journal.pone.0118415. PMC 4348419. PMID 25734540.
- Greuet, C (1968). "Organisation ultrastructurale de l'ocelle de deux Peridiniens Warnowiidae, Erythropsis pavillardi Kofoid et Swezy et Warnowia pulchra Schiller". Protistologica. 4: 209–230.
- Gregory S. Gavelis, Shiho Hayakawa, Richard A. White III, Takashi Gojobori, Curtis A. Suttle, Patrick J. Keeling, Brian S. Leander (2015). "Eye-like ocelloids are built from different endosymbiotically acquired components". Nature. 523 (7559): 204–7. Bibcode:2015Natur.523..204G. doi:10.1038/nature14593. hdl:10754/566109. PMID 26131935. S2CID 4462376. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2018-04-12. สืบค้นเมื่อ 2022-10-12.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - Oakley, Todd H.; Speiser, Daniel I. (2015). "How Complexity Originates: The Evolution of Animal Eyes". Annual Review of Ecology, Evolution, and Systematics. 46: 237–260. doi:10.1146/annurev-ecolsys-110512-135907.
- Ed Young; photographs by David Liittschwager (February 2016). "Inside the Eye: Nature's Most Exquisite Creation". National Geographic. 229 (2): 30–57.
| วิวัฒนาการ | | |
|---|---|---|
| พันธุศาสตร์ประชากร | ||
| พัฒนาการ | ||
| วิวัฒนาการของอนุกรมวิธาน | ||
| วิวัฒนาการของอวัยวะ | ||
| วิวัฒนาการของชีวกระบวนการ | ||
| กระแสชีวิตและแบบจำลอง | ||
| แบบจำลองสายพันธุ์ใหม่ | ||
| บทความที่เกี่ยวข้อง | ||
| ประวัติชีวศาสตร์ | ||
- CS1 maint: uses authors parameter
- บทความที่ขาดแหล่งอ้างอิงเฉพาะส่วนตั้งแต่2011-05
- หน้าทั้งหมดที่ต้องการเก็บกวาด
- บทความที่ต้องการการขยายความ ตั้งแต่ 2008-12
- บทความที่ขาดแหล่งอ้างอิงเฉพาะส่วนตั้งแต่2009-04
- บทความที่ต้องการตรวจสอบความถูกต้องตั้งแต่2009-02
- บทความที่ต้องการการขยายความ ตั้งแต่ 2017-03
- บทความที่ขาดแหล่งอ้างอิงเฉพาะส่วนตั้งแต่2016-03
- บทความที่มีลิงก์เสียตั้งแต่November 2017
- วิวัฒนาการ
- วิวัฒนาการโดยลักษณะปรากฏ
- ตา