
หูชั้นในรูปหอยโข่ง
| หูชั้นในรูปหอยโข่ง (Cochlea) | |
|---|---|
 ภาพตัดขวางของหูชั้นในรูปหอยโข่ง | |
| รายละเอียด | |
| ระบบ | ระบบการได้ยิน |
| ตัวระบุ | |
| ภาษาละติน | Cochlea |
| MeSH | D003051 |
| นิวโรเล็กซ์ ID | birnlex_1190 |
| TA98 | A15.3.03.025 |
| TA2 | 6964 |
| FMA | 60201 |
| อภิธานศัพท์กายวิภาคศาสตร์ | |
หูชั้นในรูปหอยโข่ง[1] หรือ อวัยวะรูปหอยโข่ง[1] หรือ คอเคลีย (อังกฤษ: cochlea, /ˈkɒk.liə/, จาก กรีกโบราณ: κοχλίας , kōhlias, แปลว่า หมุนเป็นวงก้นหอย หรือเปลือกหอยทาก) เป็นอวัยวะรับเสียงในหูชั้นใน เป็นช่องกลวงมีรูปร่างเป็นก้นหอยโข่งอยู่ในกระดูกห้องหูชั้นใน (bony labyrinth) โดยในมนุษย์จะหมุน 2.5 ครั้งรอบ ๆ แกนที่เรียกว่า modiolus[2][3] และมีเส้นผ่าศูนย์กลางประมาณ 9 มม.[4] โครงสร้างหลักของคอเคลียก็คืออวัยวะของคอร์ติ ซึ่งเป็นอวัยวะรับประสาทสัมผัสคือการได้ยินเสียง และอยู่กระจายไปตามผนังที่กั้นโพรงสองโพรงที่เต็มไปด้วยน้ำ ซึ่งวิ่งไปตามก้นหอยที่ค่อย ๆ แคบลง ส่วนคำว่าคอเคลียมาจากคำในภาษาละตินซึ่งแปลได้ว่า "เปลือกหอยทาก" และก็มาจากคำภาษากรีกว่า κοχλίας, kokhlias ซึ่งแปลว่า หอยทาก หรือเกลียว ซึ่งมาจากคำว่า κόχλος, kokhlos ซึ่งแปลว่า เปลือกวนเป็นก้นหอย[5] คอเคลียในสัตว์เลี้ยงลูกด้วยนมทั้งหมดยกเว้นของโมโนทรีมมีรูปร่างเช่นนี้

โครงสร้าง
[แก้]คอเคลียเป็นโพรงกระดูกรูปกรวยวนเป็นรูปก้นหอย ที่คลื่นความดันเสียงจะแพร่กระจายไปจากส่วนฐาน (base) ใกล้หูชั้นกลางที่ช่องรูปไข่ (oval window) ไปยังส่วนยอด (apex) คือที่ส่วนปลายหรือตรงกลางของก้นหอย โพรงที่วนเป็นก้นหอยเรียกว่า Rosenthal's canal หรือ spiral canal of the cochlea โดยยาวประมาณ 30 มม. และหมุน 2¾ รอบ ๆ แกนที่เรียกว่า modiolus โดยมีโครงสร้างรวมทั้ง
- มีโพรง 3 โพรงที่เรียกว่า scalae
- vestibular duct หรือ scala vestibuli บรรจุน้ำที่เรียกว่า perilymph และอยู่ด้านบนของโพรงกระดูกและชิดกับช่องรูปไข่
- tympanic duct หรือ scala tympani บรรจุ perilymph และอยู่ด้านล่างของโพรงกระดูกและไปสุดที่ช่องรูปกลม (round window)
- cochlear duct/partition หรือ scala media บรรจุน้ำที่เรียกว่า endolymph เป็นเขตที่ไอออนโพแทสเซียมมีความเข้มข้นสูง และที่มัดขนที่เรียกว่า stereocilia ของเซลล์ขน (hair cell) จะจุ่มอยู่
- helicotrema เป็นจุดที่ tympanic duct และ vestibular duct มาบรรจบกันที่ยอด (apex) ของคอเคลีย
- เยื่อ Reissner's membrane ซึ่งแยก vestibular duct จาก cochlear duct
- เยื่อฐาน (basilar membrane) เป็นโครงสร้างหลักที่แยก cochlear duct จาก tympanic duct และเป็นตัวกำหนดคุณสมบัติกระจายคลื่นกลของส่วนกั้นคอเคลีย
- อวัยวะของคอร์ติ เป็นเยื่อบุผิวรับเสียง (sensory epithelium) คือชั้นของเซลล์ที่บุเยื่อฐาน เป็นเซลล์ขนที่ได้พลังงานจากความต่างศักย์ของ perilymph และ endolymph
- เซลล์ขน (hair cell) ซึ่งเป็นเซลล์รับเสียงในอวัยวะของคอร์ติ โดยมียอดคล้ายขนที่เรียกว่า stereocilia
คอเคลียเป็นส่วนของหูชั้นในที่มีรูปเหมือนกับเปลือกหอยทาก เป็นตัวรับเสียงโดยความสั่นสะเทือน ซึ่งเป็นเหตุให้ stereocilia เคลื่อน ที่ถ่ายโอนแรงสั่นสะเทือนเป็นอิมพัลส์ประสาทซึ่งส่งไปยังสมองเพื่อตีความ มีโพรง 3 โพรงที่เต็มไปด้วยน้ำ โดยโพรงที่ 3 มีอวัยวะของคอร์ติ ซึ่งสามารถตรวจจับแรงดันแล้วส่งสัญญาณตามประสาทหู (auditory nerve) ไปยังสมอง ส่วนโพรงสองโพรงนั้นเรียกว่า vestibular canal และ tympanic canal
รายละเอียดทางกายวิภาค
[แก้]คอเคลียซึ่งกลวงมีผนังทำด้วยกระดูก และมีเยื่อบุผิว (epithelium) บาง ๆ ท่อที่ขดเป็นก้นหอยนี้จะแบ่งตามยาวเกือบทั้งหมดด้วยเยื่อแบ่งส่วน ส่วนนอกสองช่อง (หรือที่เรียกว่า duct หรือ scalae) เกิดจากเยื่อแบ่งส่วนนี้ น้ำที่ตอนแรกวิ่งเข้าจากช่องรูปไข่ จะวิ่งย้อนกลับที่ยอดของท่อก้นหอย เป็นจุดที่เปลี่ยนจาก vestibular duct เป็น tympanic duct และเป็นจุดที่เรียกว่า helicotrema เพราะท่อทั้งสองเชื่อมกันที่จุดนี้ น้ำที่ดันเข้าไปใน vestibular duct จะวิ่งย้อนทางกลับมาใน tympanic duct จนกระทั่งไปดันออกที่ช่องรูปกลม ซึ่งเป็นจุดระบายความดัน เพราะว่า น้ำที่ว่านี้เกือบบีบอัดไม่ได้และผนังของคอเคลียก็เป็นกระดูกแข็ง
เยื่อแบ่งส่วนที่วิ่งไปตามคอเคลียเกือบทั้งหมดตัวเองก็เป็นท่อเต็มไปด้วยน้ำ ซึ่งเรียกว่าเป็นท่อที่ 3 เป็นช่องตรงกลางที่เรียกว่า cochlear duct น้ำในช่องที่เรียกว่า endolymph แม้จะมีอิเล็กโทรไลต์และโปรตีนเช่นกัน แต่ก็สมบูรณ์ไปด้วยไอออนโพแทสเซียม ซึ่งทำให้มีความต่างทางเคมีจากน้ำในอีก 2 ช่องที่เรียกว่า perilymph ซึ่งสมบูรณ์ไปด้วยไอออนโซเดียม ความต่างกันนี้ทำให้เกิดความต่างศักย์ไฟฟ้า
เซลล์ขนจะตั้งเรียงเป็น 4 แถวอยู่ในอวัยวะของคอร์ติตลอดความยาวของคอเคลีย 3 แถวเป็นเซลล์ขนด้านนอก (outer hair cell, OHC) และ 1 แถวเป็นเซลล์ขนด้านใน (inner hair cell, IHC) IHC เป็นตัวส่งสัญญาณออกหลักของคอเคลีย เทียบกับ OHC ที่โดยหลักได้รับสัญญาณจากสมอง ซึ่งมีอิทธิพลต่อการเคลื่อนที่ได้ (motility) ของเซลล์ โดยทำหน้าที่เป็น pre-amplifier (ตัวขยายเสียง) เชิงกลของคอเคลีย สัญญาณที่ส่งไปยัง OHC มาจากเขต olivary body ในสมอง โดยส่งผ่านมัดใยประสาท medial olivocochlear bundle
ตัว cochlear duct ซับซ้อนแทบเท่ากับหูเอง โดยล้อมปิดสามด้านด้วยเยื่อฐาน (basilar membrane), stria vascularis, และ Reissner's membrane ส่วน Stria vascularis จะเป็นจุดที่สมบูรณ์ไปด้วยหลอดเลือดฝอยและเซลล์หลั่ง (secretory cell) ส่วน Reissner's membrane เป็นเยื่อบาง ๆ ที่แยก endolymph จาก perilymph และเยื่อฐาน เป็นส่วนที่ค่อนข้างแข็ง เป็นที่อยู่ของประสาทสัมผัสในการได้ยิน คือ อวัยวะของคอร์ติ และเป็นตัวกำหนดคุณสมบัติการกระจายคลื่นกลของคอเคลีย
หน้าที่
[แก้]คอเคลียเต็มไปด้วยน้ำที่เรียกว่า perilymph ซึ่งไหวไปตามแรงสั่นสะเทือนที่มาจากหูชั้นกลางโดยผ่านช่องรูปไข่ (oval window) เมื่อน้ำในคอเคลียเคลื่อน ท่อคอเคลีย (cochlear duct/partition) ที่อยู่ตรงกลาง ซึ่งรวมเยื่อฐาน (basilar membrane) และอวัยวะของคอร์ติ ก็จะเคลื่อนด้วย เซลล์ขนนับเป็นพัน ๆ จะรับรู้การเคลื่อนไหวผ่าน stereocilia แล้วแปลงแรงกลนั้นให้เป็นสัญญาณไฟฟ้าที่สื่อประสาทไปยังเซลล์ประสาทเป็นพัน ๆ โดยตัวนิวรอนที่ต่อกับเซลล์ขนจะเป็นตัวแปลงสัญญาณจากเซลล์ขนให้เป็นอิมพัลส์ไฟฟ้าประสาทที่เรียกว่า ศักยะงาน (action potential) ซึ่งส่งทางเส้นประสาท auditory nerve ไปยังโครงสร้างในก้านสมองเพื่อการประมวลผลต่อ ๆ ไป
รายละเอียด
[แก้]กระดูกเล็ก (ossicle) ที่เรียกว่ากระดูกโกลน เป็นตัวส่งแรงสั่นสะเทือนจากหูชั้นกลางไปยัง fenestra ovalis หรือช่องรูปไข่ ที่นอกคอเคลีย ซึ่งก็จะสั่นน้ำ perilymph ใน vestibular duct คือช่องบนของคอเคลีย กระดูกเล็ก 3 ชิ้นในหูชั้นกลางจำเป็นเพื่อส่งคลื่นเสียงเข้าไปในคอเคลีย เพราะว่าคอเคลียเป็นระบบน้ำ-เยื่อ และมันจำเป็นต้องได้แรงดันเพื่อส่งเสียงเป็นคลื่นน้ำสูงกว่าที่เป็นคลื่นอากาศโดยต่างจากหูชั้นนอก เป็นความดันที่เพิ่มขึ้นอาศัยอัตราพื้นที่ของแก้วหู (tympanic membrane) ต่อของเยื่อช่องรูปไข่ เป็นประมาณ 20 เท่า ของแรงดันคลื่นเสียงเดิมในอากาศ กระบวนการเพิ่มความดันแบบนี้เป็นรูปแบบหนึ่งของ impedance matching คือเป็นการจับคู่คลื่นเสียงที่วิ่งไปในอากาศให้เข้ากับคลื่นเสียงที่วิ่งไปในระบบน้ำ-เยื่อ
ที่ฐานของคอเคลีย duct แต่ละท่อจะจบลงที่หน้าต่างเนื้อเยื่อที่ชนกับช่องหูชั้นกลาง โดย vestibular duct จะจบที่ช่องรูปไข่ ซึ่งชนกับฐานของกระดูกโกลน เป็นจุดที่ฐานกระดูกโกลนจะสั่นเพื่อส่งแรงดันต่อจากกระดูกเล็ก และคลื่นใน perilymph ก็จะวิ่งไปจากฐานกระดูกโกลนไปทาง helicotrema แต่เพราะคลื่นน้ำจะขยับส่วนกั้นคอเคลีย (cochlear duct/partition) ขึ้นและลง ดังนั้นคลื่นที่วิ่งไปใน vestibular duct จะมีส่วนเท่ากับคลื่นที่วิ่งกลับมาใน tympanic duct ซึ่งไปสุดที่เยื่อช่องรูปกลม ทำให้เยื่อนูนออกไปทางหูชั้นกลาง เทียบกับขณะที่เมื่อเสียงวิ่งผ่านเยื่อช่องรูปไข่ ซึ่งเยื่อจะนูนเข้ามาในคอเคลีย perilymph ใน vestibular duct และ endolymph ใน cochlear duct จะมีพฤติกรรมเหมือนกับอยู่เป็นท่อเดียวกัน โดยแยกจากกันด้วยเยื่อ Reissner's membrane บาง ๆ เท่านั้น
แรงสะเทือนของ endolymph ใน cochlear duct จะเคลื่อนเยื่อฐานมากที่สุด (peak) ตามระยะจากช่องรูปไข่ขึ้นอยู่กับความถี่เสียง OHC จะทำให้อวัยวะของคอร์ติสั่นซึ่งขยายแรงดันนี้เพิ่มขึ้น IHC ก็จะเคลื่อนไปตามแรงสั่นที่วิ่งไปตามน้ำ และเกิดการลดขั้ว (depolarization) เมื่อไอออน K+ วิ่งเข้าช่องไอออนที่เปิดปิดโดยใยเชื่อมปลายขน (tip-link) และส่งสัญญาณโดยใช้สารสื่อประสาทไปยังนิวรอนของ spiral ganglion (ซึ่งเป็นตัวส่งสัญญาณต่อไปยังสมอง) เซลล์ขนในอวัยวะของคอร์ติจะเลือกตัวกระตุ้นเป็นความถี่เสียงโดยเฉพาะขึ้นอยู่กับตำแหน่งในคอเคลีย ตามความอ่อนแข็งของ basilar membrane[6]
ความอ่อนแข็งจะขึ้นอยู่กับปัจจัยหลายอย่างรวมทั้งความหนาและความกว้างของเยื่อ[7] ซึ่งแข็งสุดตรงจุดใกล้กับช่องรูปไข่ที่สุด ที่กระดูกโกลนส่งความสั่นสะเทือนต่อมาจากแก้วหู เนื่องจากแข็งที่สุดตรงนี้ จึงจะขยับเหตุความสั่นความถี่สูงเท่านั้น โดยเซลล์ขนจะขยับไปพร้อม ๆ กัน คลื่นยิ่งเดินเข้าไปทางยอดของคอเคลียคือจุด helicotrema เท่าไร เยื่อฐานก็จะแข็งน้อยลงเท่านั้น ดังนั้น คลื่นความถี่ต่ำจะดำเนินยาวไปในท่อ ไปสู่จุดที่เยื่อแข็งน้อยกว่าและขยับได้ง่ายกว่า คือ เมื่อคลื่นดำเนินไปสู่ที่อ่อนกว่า คลื่นจะวิ่งช้าลงโดยตัวเยื่อจะตอบสนองต่อคลื่นได้ดีกว่า นอกจากนั้นแล้ว ในสัตว์เลี้ยงลูกด้วยนม คอเคลียจะม้วนเป็นก้นหอย ซึ่งมีหลักฐานว่าช่วยเพิ่มแรงสั่นสะเทือนความถี่ต่ำเมื่อคลื่นวิ่งไปตามก้นหอยที่เต็มไปด้วยน้ำ[8]
การเรียงลำดับความถี่การรู้เสียงตามปริภูมิเช่นนี้เรียกว่า tonotopy สำหรับเสียงที่ต่ำมาก (คือน้อยกว่า 20 Hz) คลื่นเสียงจะเดินไปจนสุดทาง คือตาม vestibular duct และ tympanic duct จนถึงจุด helicotrema เสียงความถี่ต่ำขนาดนี้ยังทำให้อวัยวะคอร์ติทำงานบ้าง แต่ว่า ก็ต่ำเกินที่จะให้ทราบความถี่ที่ชัดเจน ส่วนเสียงความถี่สูงจะไม่เดินไปถึงจุด helicotrema เนื่องจากการเรียงลำดับความแข็งแบบ tonotopy การเคลื่อนไหวของเยื่อฐานแบบรุนแรงเนื่องจากเสียงดังอาจทำให้เซลล์ขนตาย นี่เป็นเหตุสามัญของการเสียการได้ยิน และเป็นเหตุที่ทำให้คนที่ใช้ปืนหรือเครื่องจักรหนักบ่อยครั้งจำเป็นต้องใส่เครื่องป้องกันหู
การขยายเสียงของเซลล์ขน
[แก้]คอเคลียไม่เพียงแต่ "รับ" เสียงเท่านั้น แต่ยัง "สร้าง" เสียงและขยายเสียงเมื่อยังมีสภาพดี เมื่อสิ่งมีชีวิตจำเป็นต้องได้ยินเสียงที่ค่อย คอเคลียก็จะขยายเสียงโดยการถ่ายโอนความรู้สึกแบบผกผัน (reverse transduction) ที่เซลล์ขนด้านนอก (OHC) คือจะเปลี่ยนสัญญาณไฟฟ้ากลับไปเป็นแรงกลผ่านกระบวนการป้อนกลับเชิงบวก โดย OHC มีมอเตอร์โปรตีนที่เรียกว่า prestin ที่เยื่อหุ้มเซลล์ด้านนอก ซึ่งสร้างความเคลื่อนไหวโดยเพิ่มแรงดันคลื่นน้ำที่มากระทบกับเยื่อหุ้มเซลล์ การขยายเสียงแบบแอคทีฟเช่นนี้ สำคัญเพื่อให้ได้ยินเสียงค่อย ๆ แต่ก็ทำให้เกิดปรากฏการณ์ที่คอเคลียสร้างคลื่นเสียงแล้วส่งย้อนกลับเข้ามาผ่านหูชั้นกลางที่เรียกว่า otoacoustic emissions
Otoacoustic emissions
[แก้]Otoacoustic emission (การปล่อยเสียงจากหู) เป็นคลื่นเสียงที่เกิดจากคอเคลียผ่านช่องรูปไข่ (oval window) กระจายต่อไปผ่านหูชั้นกลางจนถึงแก้วหู ออกจากรูหูแล้วสามารถเก็บเสียงได้โดยไมโครโฟน การปล่อยเสียงจากหูสำคัญในการทดสอบความพิการในการได้ยินบางอย่าง เพราะว่า เสียงนี้จะมีก็ต่อเมื่อคอเคลียอยู่ในสภาพดี และจะมีน้อยลงถ้า OHC ทำงานลดลง
บทบาทของแกบจังก์ชัน
[แก้]โปรตีนแกบจังก์ชันชื่อว่า connexin ที่พบในคอเคลียมีบทบาทสำคัญต่อการได้ยิน[9] การกลายพันธุ์ของยีนประเภทนี้มีหลักฐานว่า ทำให้เกิดหูหนวกทั้งแบบมีอาการและแบบไม่มีอาการ[10] connexin บางอย่าง รวมทั้ง connexin 30 (GJB6) และ connexin 26 (GJB2) มีอย่างแพร่หลายในระบบแกบจังก์ชันสองอย่างที่พบเฉพาะในคอเคลีย เครือข่าย epithelial-cell gap-junction จะจับคู่เซลล์บุผิวที่ไม่เกี่ยวกับการได้ยิน (non-sensory epithelial cell) ในขณะที่เครือข่าย connective-tissue gap-junction จะจับคู่เซลล์เนื้อเยื่อเกี่ยวพัน (connective-tissue cell) โดยช่องแกบจังก์ชันจะทำหน้าที่เวียนไอออนโพแทสเซียมกลับไปที่ endolymph หลังจากการถ่ายโอนแรงกล (mechanotransduction) ของเซลล์ขน[11] จุดที่สำคัญก็คือ แกบจังก์ชันจะพบในระหว่างเซลล์สนับสนุนของคอเคลีย แต่ไม่พบในเซลล์ขนที่ทำให้ได้ยิน[12]
ความสำคัญทางคลินิก
[แก้]ชีวประดิษฐ์
[แก้]ในปี 2552 วิศวกรที่สถาบันเทคโนโลยีแมสซาชูเซตส์สร้างวงจรเบ็ดเสร็จเป็นชิปที่สามารถวิเคราะห์ความถี่คลื่นวิทยุในพิสัยกว้างมาก แต่กินไฟเพียงแค่ส่วนหนึ่งของเทคโนโลยีที่มีอยู่แล้ว โดยเลียนแบบการทำงานของคอเคลีย[13][14]
ในสัตว์อื่น
[แก้]คอเคลียในรูปแบบก้นหอยพบแต่ในสัตว์เลี้ยงลูกด้วยนมเท่านั้น ส่วนในสัตว์ปีกและสัตว์มีกระดูกสันหลังอื่นที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนม ช่องที่มีเซลล์รับเสียงบ่อยครั้งก็เรียกว่า คอเคลีย เหมือนกัน แม้ว่าจะไม่เป็นรูปหอยโข่ง แต่จะเป็นท่อตันที่เรียกว่า cochlear duct
ความแตกต่างเช่นนี้ดูเหมือนจะวิวัฒนาการไปพร้อม ๆ กับความแตกต่างในการได้ยินพิสัยความถี่เสียงในระหว่างสัตว์เลี้ยงลูกด้วยนมและสัตว์มีกระดูกสันหลังอื่น คือสัตว์เลี้ยงลูกด้วยนมมีพิสัยความถี่เสียงที่กว้างกว่า โดยส่วนหนึ่งก็เพราะกลไกโดยเฉพาะในการขยายเสียง (pre-amplification) ที่ใช้การสั่นตัวของเซลล์ขนด้านนอก (OHC) แต่ความละเอียดของการได้ยินความถี่เสียงไม่ได้ดีกว่าในสัตว์เลี้ยงลูกด้วยนมเทียบกับสัตว์เลื้อยคลานและสัตว์ปีกโดยมาก เพียงแต่ว่า ความถี่เสียงสูงสุดที่ได้ยินมักจะสูงกว่า และบางครั้งจะสูงกว่ามาก
สปีชีส์นกโดยมากไม่ได้ยินเสียงสูงกว่า 4-5 kHz โดยนกแสกได้ยินเสียงสูงสุดอยู่ที่ ~11 kHz แต่มีสัตว์เลี้ยงลูกด้วยนมน้ำบางชนิดที่ได้ยินเสียงสูงถึง 200 kHz การมีช่องที่ขดเป็นก้นหอย แทนที่จะเป็นช่องตรง ๆ สั้น ๆ ทำให้มีเนื้อที่สำหรับได้ยินเสียงในพิสัยความถี่ที่กว้างกว่า ทำให้สัตว์เลี้ยงลูกด้วยนมสามารถมีพฤติกรรมบางอย่างโดยเฉพาะเกี่ยวกับการได้ยินเสียง[15]
เพราะว่าการศึกษาคอเคลียควรจะเล็งที่ระดับเซลล์ขน การรู้ความแตกต่างทางกายวิภาคและสรีรภาพของเซลล์ขนในสปีชีส์ต่าง ๆ จึงสำคัญ ตัวอย่างเช่น สัตว์ปีก แทนที่จะมีเซลล์ขนด้านนอกและเซลล์ขนด้านใน ก็มีเซลล์ขนยาว (tall hair cell) และเซลล์ขนสั้น (short hair cell) และก็มีอะไรอย่างอื่น ๆ ที่ดูจะเหมือนกัน ยกตัวอย่างเช่น เซลล์ขนยาวมีหน้าที่คล้ายกับเซลล์ขนด้านใน และเซลล์ขนสั้น เพราะไม่มีใยประสาทที่ส่งไปยังระบบประสาทกลาง ก็จึงเหมือนกับเซลล์ขนด้านนอก และตัวอย่างความแตกต่างก็คือ เซลล์ขนทั้งหมดจะอยู่ติดกับ tectorial membrane ในสัตว์ปีก เทียบกับสัตว์เลี้ยงลูกด้วยนม ที่เซลล์ขนด้านนอกเท่านั้นที่อยู่ติดกับ tectorial membrane
รูปภาพอื่น ๆ
[แก้]-
 ภาพ MRI ของคอเคลียและหลอดครึ่งวงกลม
ภาพ MRI ของคอเคลียและหลอดครึ่งวงกลม -
 ห้องหูชั้นใน
ห้องหูชั้นใน -
 กระดูกห้องหูชั้นใน (bony labyrinth) ด้านขวา มองจากด้านข้าง
กระดูกห้องหูชั้นใน (bony labyrinth) ด้านขวา มองจากด้านข้าง -
 ข้างในกระดูกห้องหูชั้นใน (bony labyrinth) ด้านขวา
ข้างในกระดูกห้องหูชั้นใน (bony labyrinth) ด้านขวา -
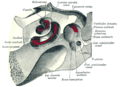 คอเคลียและโพรงหู มองจากด้านบน
คอเคลียและโพรงหู มองจากด้านบน -
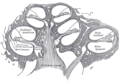 ผังคอเคลียผ่าตามยาว (longitudinal section)
ผังคอเคลียผ่าตามยาว (longitudinal section)






ดูเพิ่ม
[แก้]เชิงอรรถและอ้างอิง
[แก้]- ↑ 1.0 1.1 "cochlea", ศัพท์บัญญัติอังกฤษ-ไทย, ไทย-อังกฤษ ฉบับราชบัณฑิตยสถาน (คอมพิวเตอร์) รุ่น ๑.๑ ฉบับ ๒๕๔๕,
(แพทยศาสตร์) ๑. อวัยวะรูปหอยโข่ง ๒. หูชั้นในรูปหอยโข่ง
- ↑
Gilroy, Anne M; MacPherson, Brian R; Ross, Lawrence M (2008). Atlas of anatomy. Thieme. p. 536. ISBN 978-1-60406-151-2.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑
Moore and Dalley (1999). Clinically Oriented Anatomy (4th ed.). p. 974.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Principles of Neural Science, 5th edition (2013), "Chapter 31: The Inner Ear", pp. 656
- ↑ "etymology of "cochleㄷa"". Online Etymology Dictionary.
- ↑ Ehret, Guenter (December 1978). "Stiffness gradient along the basilar membrane as a way for spatial frequency analysis within the cochlea" (PDF). J Acoust Soc Am. 64 (6): 1723–6. doi:10.1121/1.382153. PMID 739099.
- ↑ Camhi, J (1984). Neuroethology: nerve cells and the natural behavior of animals. Sinauer Associates.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Manoussaki, D; Chadwick, RS; Ketten, DR; Arruda, J; Dimitriadis, EK; O'Malley, JT (2008). "The influence of cochlear shape on low-frequency hearing". Proc Natl Acad Sci U S A. 105 (16): 6162–6166. doi:10.1073/pnas.0710037105. PMC 2299218. PMID 18413615.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Zhao, H. -B.; Kikuchi, T.; Ngezahayo, A.; White, T. W. (2006). "Gap Junctions and Cochlear Homeostasis". Journal of Membrane Biology. 209 (2–3): 177–186. doi:10.1007/s00232-005-0832-x. PMC 1609193. PMID 16773501.
- ↑ Erbe, C. B.; Harris, K. C.; Runge-Samuelson, C. L.; Flanary, V. A.; Wackym, P. A. (2004). "Connexin 26 and Connexin 30 Mutations in Children with Nonsyndromic Hearing Loss". The Laryngoscope. 114 (4): 607–611. doi:10.1097/00005537-200404000-00003. PMID 15064611.
- ↑ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Takasaka, T.; Adams, J. C. (2000). "Gap junction systems in the mammalian cochlea". Brain research. Brain research reviews. 32 (1): 163–166. doi:10.1016/S0165-0173(99)00076-4. PMID 10751665.
- ↑ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Adams, J. C. (1995). "Gap junctions in the rat cochlea: Immunohistochemical and ultrastructural analysis". Anatomy and embryology. 191 (2): 101–118. doi:10.1007/BF00186783. PMID 7726389.
- ↑
Trafton, Anne (3 June 2009). MIT newsoffice http://web.mit.edu/newsoffice/2009/bio-electronics-0603.html.
{{cite web}}:|title=ไม่มีหรือว่างเปล่า (help); ไม่รู้จักพารามิเตอร์|___title=ถูกละเว้น (help) - ↑
Mandal, Soumyajit; Zhak, Serhii M; Sarpeshkar, Rahul (June 2009). "A Bio-Inspired Active Radio-Frequency Silicon Cochlea". IEEE Journal of Solid-State Circuits. 44 (6): 1814–1828. doi:10.1109/JSSC.2009.2020465.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Vater, M; Meng, J; Fox, RC (2004). Hearing organ evolution and specialization: Early and later mammals. Evolution of the Vertebrate Auditory System. New York: Springer-Verlag. pp. 256–288.
{{cite book}}: ไม่รู้จักพารามิเตอร์|editors=ถูกละเว้น แนะนำ (|editor=) (help)CS1 maint: uses authors parameter (ลิงก์)
- Kandel, Eric R; Schwartz, James H; Jessell, Thomas M; Siegelbaum, Steven A; Hudspeth, AJ (2013). Principles of Neural Science (5th ed.). United State of America: McGraw-Hill. ISBN 978-0-07-139011-8.
- Physiology of the Ear By Anthony F. Jahn, Joseph Santos-Sacchi
- Audiology By Ross J. Roeser, Michael Valente, Holly Hosford-Dunn
- Audition By Michel Imbert, R. H. Kay
- The Cochlea By Peter Dallos, Arthur N. Popper, Richard R. Fay
แหล่งข้อมูลอื่น
[แก้]- Cochlea ในหอสมุดแพทยศาสตร์แห่งชาติอเมริกัน สำหรับหัวข้อเนื้อหาทางการแพทย์ (MeSH)
- "Promenade 'Round the Cochlea" by R. Pujol, S. Blatrix, T. Pujol et al. at University of Montpellier
- "Histology Videos of The Ear"