
ผลต่างระหว่างรุ่นของ "สฟิงโกไมอีลิน"
ไม่มีความย่อการแก้ไข |
ไม่มีความย่อการแก้ไข |
||
| บรรทัด 28: | บรรทัด 28: | ||
สฟิงโกไมอีลินมีปฏิกิริยาสำคัญกับคอเลสเตอรอล [[คอเลสเตอรอล]]มีคุณสมบัติในการหยุดการเปลี่ยนสถานะจากของเหลวเป็นของแข็งในฟอสโฟลิพิด เนื่องจากอุณหภูมิในการเปลี่ยนสถานะของสฟิงโกไมอีลินอยู่ในช่วงอุณหภูมิสรีรวิทยาของร่างกาย คอเลสเตอรอลจึงสามารถมีบทบาทสำคัญในสถานะของสฟิงโกไมอีลิน สฟิงโกไมอีลินยังมีแนวโน้มสร้างพันธะไฮโดรเจนกับโมเลกุลอื่นมากกว่าฟอสโฟลิพิดชนิดอื่น<ref>Massey JB. Interaction of ceramides with phosphatidylcholine, sphingomyelin and sphingomyelin/cholesterol bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2001 2/9;1510 (1–2) :167-84.</ref> |
สฟิงโกไมอีลินมีปฏิกิริยาสำคัญกับคอเลสเตอรอล [[คอเลสเตอรอล]]มีคุณสมบัติในการหยุดการเปลี่ยนสถานะจากของเหลวเป็นของแข็งในฟอสโฟลิพิด เนื่องจากอุณหภูมิในการเปลี่ยนสถานะของสฟิงโกไมอีลินอยู่ในช่วงอุณหภูมิสรีรวิทยาของร่างกาย คอเลสเตอรอลจึงสามารถมีบทบาทสำคัญในสถานะของสฟิงโกไมอีลิน สฟิงโกไมอีลินยังมีแนวโน้มสร้างพันธะไฮโดรเจนกับโมเลกุลอื่นมากกว่าฟอสโฟลิพิดชนิดอื่น<ref>Massey JB. Interaction of ceramides with phosphatidylcholine, sphingomyelin and sphingomyelin/cholesterol bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2001 2/9;1510 (1–2) :167-84.</ref> |
||
=== ตำแหน่ง === |
|||
สฟิงโกไมอีลินถูกสังเคราะห์ที่[[ร่างแหเอนโดพลาซึม]] (endoplasmic reticulum) และ ทรานส์-[[กอลจิคอมเพล็กซ์]] พบมากใน[[เยื่อหุ้มเซลล์]]โดยมีความเข้มข้นมากในเนื้อเยื่อชั้นนอกมากกว่าชั้นใน<ref name="Testi"/> The Golgi complex represents an intermediate between the ER and plasma membrane, with slightly higher concentrations towards the trans side.<ref>Bragger B, Sandhoff R, Wegehingel S, Gorgas K, Malsam J, Helms JB, Lehmann W, Nickel W, Wieland FT. Evidence for segregation of sphingomyelin and cholesterol during formation of COPI-coated vesicles. J Cell Biol 2000 10/30;151 (3) :507-17</ref> |
|||
== ภาพอื่นๆ == |
== ภาพอื่นๆ == |
||
รุ่นแก้ไขเมื่อ 00:37, 20 พฤษภาคม 2561

สฟิงโกไมอีลิน (อังกฤษ: sphingomyelin) เป็นสฟิงโกลิพิดชนิดหนึ่งที่พบในเยื่อหุ้มเซลล์ของสัตว์ โดยเฉพาะเยื่อไมอีลินที่หุ้มรอบแอกซอนของเซลล์ประสาท ส่วนใหญ่ประกอบด้วยฟอสโฟโคลีน (phosphocholine) และเซราไมด์ (ceramide) หรือหมู่ฟังก์ชันเอทาโนลามีน (ethanolamine) ดังนั้นสฟิงโกไมอีลินจึงสามารถจัดอยู่ในกลุ่มสฟิงโกฟอสโฟลิพิด (sphingophospholipids)[1] ในมนุษย์ สฟิงโกไมอีลินนับเป็นประมาณร้อยละ 85 ของสฟิงโกลิพิดในร่างกาย และคิดเป็น 10–20 ร้อยละโมลของลิพิดในเยื่อหุ้มเซลล์
สฟิงโกไมอีลินถูกแยกครั้งแรกโดยนักเคมีชาวเยอรมัน Johann L.W. Thudicum ในทศวรรษที่ 1880[2] โครงสร้างของสฟิงโกไมอีลินถูกรายงานครั้งแรกในปี ค.ศ. 1927 ในชื่อ เอ็น-เอซิล-สฟิงโกซีน-1-ฟอสฟอริลโคลีน (N-acyl-sphingosine-1-phosphorylcholine)[2] ในสัตว์เลี้ยงลูกด้วยนม สฟิงโกไมอีลินเป็นส่วนประกอบตั้งแต่ร้อยละ 2-15 ในเนื้อเยื่อส่วนใหญ่โดยพบมากในเนื้อเยื่อประสาท เม็ดเลือดแดง และในเลนส์ตา สฟิงโกไมอีลินมีบทบาทสำคัญในโครงสร้างและการทำงานของเซลล์ เป็นส่วนประกอบของเยื่อหุ้มเซลล์และมีส่วนในการถ่ายทองสัญญาณ เมแทบอลิซึมของสฟิงโกไมอีลินทำให้เกิดผลิตภัณฑ์ที่มีส่วนสำคัญภายในเซลล์[2]
โมเลกุลสฟิงโกไมอีลินประกอบด้วยฟอสโฟโคลีน (phosphocholine) หรือฟอสโฟเอทานอลามีน (phosphoethanolamine) เป็นหมู่ฟังก์ชันส่วนหัวที่มีขั้ว และจัดเป็นฟอสโฟลิพิดเช่นเดียวกับกลีเซรอฟอสโฟลิพิด (glycerophospholipid) สฟิงโกไมอีลินมีความคล้ายฟอลฟาทิดิลโคลีน (phosphatidylcholine) ในแง่คุณสมบัติทั่วไปและโครงสร้างสามมิติรวมทั้งไม่มีขั้วสุทธิในส่วนหัว เนื่องจากสฟิงโกไมอีลินพบในเยื่อหุ้มเซลล์ของเซลล์สัตว์โดยเฉพาะอย่างยิ่งในเยื่อไมอีลินที่หุ้มและเป็นฉนวนของแอกซอนในเซลล์ประสาท จึงมีชื่อว่า สฟิงโกไมอีลิน[1]
ลักษณะทางกายภาพ

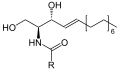
ดำ: สฟิงโกซีน; แดง: ฟอสโฟโคลีน; น้ำเงิน: กรดไขมัน
องค์ประกอบ
โครงสร้างของสฟิงโกไมอีลินประกอบด้วยหมู่ฟังก์ชันส่วนหัวฟอสโฟโคลีน (phosphocholine) สฟิงโกซีน (sphingosine) และกรดไขมัน มันเป็นหนึ่งในฟอสโฟลิพิดในเยื่อหุ้มส่วนน้อยที่ไม่ได้สังเคราะห์มาจากกลีเซอรอล สฟิงโกซีนและกรดไขมันสามารถเรียกรวมกันว่าเซราไมด์ (ceramide) องค์ประกอบดังกล่าวทำให้สฟิงโกไมอีลินมีบทบาทสำคัญในวิถีการส่งสัญญาณระดับเซลล์ กล่าวคือการย่อยสลายและสังเคราะห์สฟิงโกไมอีลินทำให้เกิดสารนำรหัสสำหรับการถ่ายโอนสัญญาณ
สฟิงโกไมอีลินพบในธรรมชาติเช่นในไข่ หรือสมองสัตว์ ซึ่งจะมีกรดไขมันที่มีความยาวหลากหลาย สฟิงโกไมอีลินที่มีความยาวจำเพาะเช่น ปาล์มิโตอิลสฟิงโกไมอีลิน (palmitoylsphingomyelin) ซึ่งมีกรดไขมันอิ่มตัว 16 หมู่เอซิลมีจำหน่ายเพื่อการค้า[3]
คุณสมบัติ
ตามหลักการแล้วโมเลกุลสฟิงโกไมอีลินมีรูปร่างคล้ายทรงกระบอก แต่โมเลกุลสฟิงโกไมอีลินจำนวนมากที่สายไฮโดรโฟบิกไม่เข้าคู่กัน คือมีความยาวระหว่างสองสายที่แตกต่างกันมาก[4] สายไฮโดรโฟบิกของสฟิงโกไมอีลินมีแนวโน้มที่จะอิ่มตัวมากกว่าฟอสโฟลิพิดชนิดอื่น อุณหภูมิในการเปลี่ยนสถานะหลักของสฟิงโกไมอีลินยังสูงกว่าเมื่อเทียบกับฟอสโฟลิพิดที่คล้ายกัน คือประมาณ 37 องศาเซลเซียส ซึ่งทำให้เกิดความแตกแบบกันในทางด้านข้างของเยื่อหุ้มและเกิดเป็นขอบเขตในเยื่อหุ้มชนิดสองชั้น[4]
สฟิงโกไมอีลินมีปฏิกิริยาสำคัญกับคอเลสเตอรอล คอเลสเตอรอลมีคุณสมบัติในการหยุดการเปลี่ยนสถานะจากของเหลวเป็นของแข็งในฟอสโฟลิพิด เนื่องจากอุณหภูมิในการเปลี่ยนสถานะของสฟิงโกไมอีลินอยู่ในช่วงอุณหภูมิสรีรวิทยาของร่างกาย คอเลสเตอรอลจึงสามารถมีบทบาทสำคัญในสถานะของสฟิงโกไมอีลิน สฟิงโกไมอีลินยังมีแนวโน้มสร้างพันธะไฮโดรเจนกับโมเลกุลอื่นมากกว่าฟอสโฟลิพิดชนิดอื่น[5]
ตำแหน่ง
สฟิงโกไมอีลินถูกสังเคราะห์ที่ร่างแหเอนโดพลาซึม (endoplasmic reticulum) และ ทรานส์-กอลจิคอมเพล็กซ์ พบมากในเยื่อหุ้มเซลล์โดยมีความเข้มข้นมากในเนื้อเยื่อชั้นนอกมากกว่าชั้นใน[6] The Golgi complex represents an intermediate between the ER and plasma membrane, with slightly higher concentrations towards the trans side.[7]
ภาพอื่นๆ
-
 แบบจำลอง ball-and-stick ของสฟิงโกไมอีลิน
แบบจำลอง ball-and-stick ของสฟิงโกไมอีลิน -
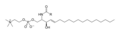 สูตรโครงสร้างแบบเส้นของสฟิงโกไมอีลิน
สูตรโครงสร้างแบบเส้นของสฟิงโกไมอีลิน -
-


อ้างอิง
- ↑ 1.0 1.1 Donald J. Voet; Judith G. Voet; Charlotte W. Pratt (2008). "Lipids, Bilayers and Membranes". Principles of Biochemistry, Third edition. Wiley. p. 252. ISBN 978-0470-23396-2.
- ↑ 2.0 2.1 2.2 Ramstedt B, Slotte JP. Membrane properties of sphingomyelins. FEBS Lett 2002 10/30;531 (1) :33-7
- ↑ "Archived copy". คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2014-03-29. สืบค้นเมื่อ 2013-07-16.
{{cite web}}: ไม่รู้จักพารามิเตอร์|deadurl=ถูกละเว้น แนะนำ (|url-status=) (help)CS1 maint: archived copy as title (ลิงก์) - ↑ 4.0 4.1 Barenholz Y, Thompson TE. Sphingomyelin: Biophysical aspects. Chem Phys Lipids 1999 11;102 (1–2) :29-34.
- ↑ Massey JB. Interaction of ceramides with phosphatidylcholine, sphingomyelin and sphingomyelin/cholesterol bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2001 2/9;1510 (1–2) :167-84.
- ↑ อ้างอิงผิดพลาด: ป้ายระบุ
<ref>ไม่ถูกต้อง ไม่มีการกำหนดข้อความสำหรับอ้างอิงชื่อTesti - ↑ Bragger B, Sandhoff R, Wegehingel S, Gorgas K, Malsam J, Helms JB, Lehmann W, Nickel W, Wieland FT. Evidence for segregation of sphingomyelin and cholesterol during formation of COPI-coated vesicles. J Cell Biol 2000 10/30;151 (3) :507-17